Mindenki másképp egyforma – folytatódik a nagy genetikai őskeresés

Előző cikkünkben részben abba ástuk bele magunkat, hogy mennyire bizonytalan, ha a mitokondriális DNS-t (mtDNS) használjuk arra, hogy rekonstruáljuk a saját családunk, vagy akár egy teljes ország népességének történetét.
Persze felvethető, hogy egyáltalán releváns-e ezeket a genetikai kapcsolatokat keresni, ha egyszer a honfoglalók kultúrájának fennmaradása – úgy, hogy genetikai állományuk teljesen feloldódott az őshonos Kárpát-medencei állományban – jól mutatja, hogy az identitás elsősorban kulturális kérdés. A késő avarkori és a jelenkori Kárpát-medencei mtDNS minták hasonlósága megerősíti, amit a hazai és környékbeli őskori leletek alapján látunk. A térség genetikai összetételét alapvetően három ősi csoport, az európai „őslakos” vadászó-gyűjtögetők, az Anatóliából érkező földművesek és a Fekete-tenger északi partjától induló gödörsíros kurgán népek keveredése az időszámítás előtti harmadik évezredben hozta létre a mai európai génállományt. Ezt nagy mértékben már nem befolyásolták a különféle hódítások és népvándorlási hullámok.1 A hódítók (rómaiak, hunok, avarok vagy magyarok) elsősorban kulturálisan írják át a térségben élő emberek identitását, a génállomány nagyfokú kicserélődésére a mai adatok alapján nem látunk példát.
Genetikai identitás
Miért is határozná meg az identitását valakinek az, ha a teljes mtDNS-én (vagy Y kromoszómáján), vagy valamelyik másik kromoszómáján pár tízezer, esetleg egymillió bázispárnyi szekvenciája honfoglaló, avar, hun, szkíta vagy római ősétől származna? Már a kérdésfelvetés is abszurd, és azt tükrözi, hogy mennyire homályos gondolatok vannak a fejekben arról, hogy miképp kódolódnak és öröklődnek egyes jellegeink: mindenre igyekszünk az iskolában megtanult Mendel-féle borsójellegek egyszerű, egygénes sémáját ráhúzni, miközben a valóság szinte mindig sokkal összetettebb.
A Mendel-féle absztrakt „öröklődő egységek” fogalmától (ezeket nevezte el később géneknek a dán biológus Wilhelm Johannsen) hosszú út és sok évtizednyi kutatás vezetett ahhoz a képhez, amivel ma rendelkezünk a genomról. Messzi vagyunk ma már a genetika múlt századi első hőskorában népszerű képtől, amely a genomot egy nyaklánchoz hasonlította, ahol mint gyöngyök a zsinegen, szorosan egymás után ülnek a gének a kromoszómán. Az élővilág genomjainak szerveződéséről radikálisan más képet mutatnak a lassan két évtizede zajló genomszekvenálási projektek, ez alól az emberi genom sem kivétel.
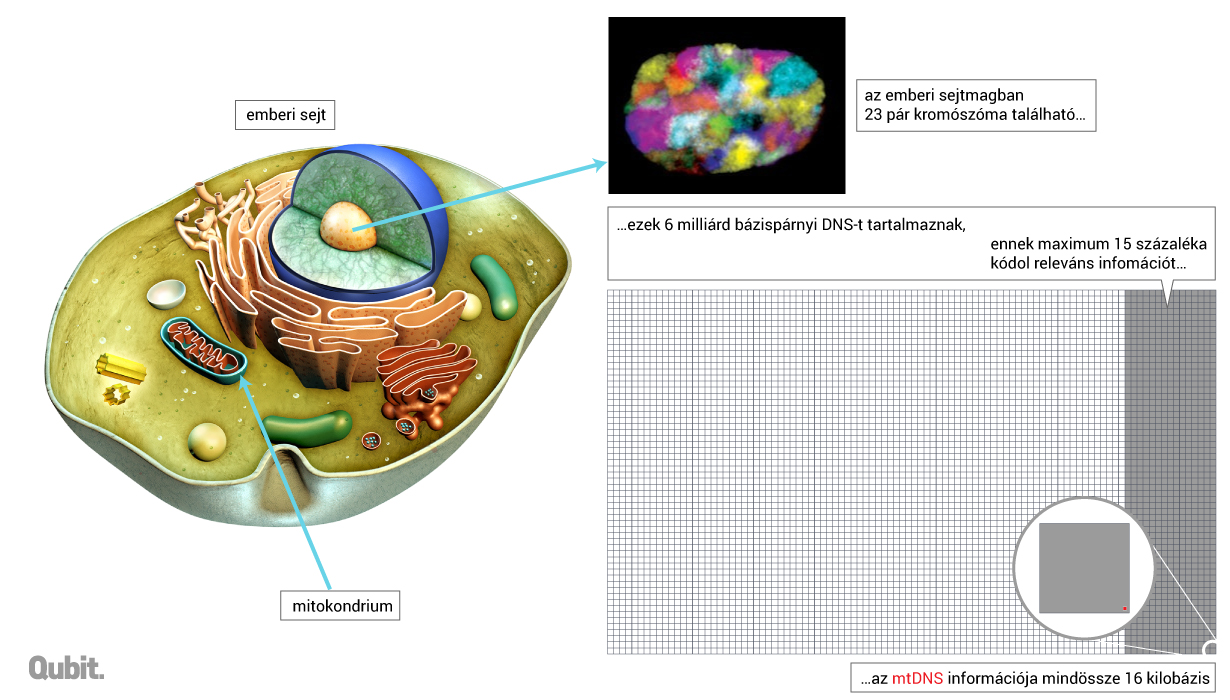
Ma már látjuk, hogy a sejt összes DNS-ének csak a töredéke hordoz olyan információt, ami valóban fontos a sejt működéséhez. Ez a legnagyvonalúbb becslések szerint is maximum a teljes örökítőanyag 15 százaléka lehet, amiben már bőven benne vannak a szabályozó szekvenciák is, hiszen a fehérjekódoló régiók hozzávetőlegesen 2 százalékát adják a genomnak.2
A genom nagyobb része nem vesz részt jellegeink kialakításában, de ugyanúgy másolódik és öröklődik, mint a nem kódoló régiók által körülvett konkrét gének. Ez azt is jelenti, hogy ugyan olykor egy sok millió bázispárt kitevő egységet örökölhetünk valamelyik felmenőnk kromoszómájából úgy, hogy közben a sejt és a szervezet működéséhez semmilyen releváns információ nem lesz kódolva. És még amikor teljes gének öröklődnek is át, azok szinte sosem biztosítják egy jelleg teljes öröklődését.3
Szemszín és Illyés Gyula
Egyszerűbb tulajdonságaink, mint a testmagasság, bőr- és hajszín is a gének és a környezet bonyolult összjátékaként jönnek létre. Még azokban az esetekben is, ahol a jellegnek egyértelműen nagyon erős genetikai meghatározottsága van, gyakran sok száz vagy ezer gén összjátéka határozza meg a végső kimenetet. Nincs például „magasság gén”, hiszen az összes kromoszómánkon szétszórt közel húszezer szekvencia együttesen határozza meg a fejlődő szervezet végső magasságát.4 Nem mind egyforma erejű, de mégis ennyi információra van szükségünk, hogy megmagyarázzuk, miképp kódolja a genom a magasságunkat. A szem színéért felelős hét gén együttes működése is még csupán a jelleg változatosságának durván háromnegyedét magyarázza meg, a fennmaradó részért valószínűleg sok száz vagy ezer kis hatású további szekvencia tehető felelőssé. Salman Rushdie legutóbbi regényének előző cikkünkben említett fülcimpás példájára visszatérve: bár sokáig valóban egyetlen géntől függő, klasszikus mendeli jellegnek tekintették a fülcimpa hiányát, alig pár hónapja derült ki, hogy a valóságban alsó hangon itt is közel ötven genomi pozíció összjátékáról van szó.5
Minél komplexebb egy tulajdonság, annál nagyobb a környezet szerepe, és annál több fehérjekódoló, illetve azok működését szabályozó DNS-darab együttes működésére van szükség kialakításához. Ez azt is jelenti, hogy a tulajdonságot létrehozó szekvenciasokaság együttes öröklődésének esélye már a közvetlen utódokban is elhanyagolható, a távolabbi leszármazottaknál pedig gyakorlatilag a nullával egyenlő. Épp ezért is egyértelmű, hogy a kulturális identitás esetében dőreség generációkon átívelő genetikai gyökereket keresni. Avar, szkíta, hun vagy honfoglaló végső soron az volt, aki annak vallotta magát. Mint ahogy magyarra sincs ennél egzaktabb definíciónk, legfeljebb Illyés Gyulát idézhetjük, aki szerint „jó magyar az, aki emberi, jó tagja a magyar közösségnek”.6
Attól, hogy tulajdonságainkban vagy gyakran genetikai állományunkban nem hordozunk semmit belőlük, persze még nem lesz kevésbé érdekes megismerni felmenőinket. Nélkülük nem lennénk itt ugyanis. De akkor vajon folyik avar vagy honfoglaló vér az ereinkben? Az öröklődés furcsa, de makacs törvényei nem csak bizonytalanságot, de bizonyosságot is teremtenek: ha csak egyetlen hunnak is maradt ma élő leszármazottja a térségben, akkor az azt is jelenti, hogy mindannyian ennek a személynek a leszármazottai (is) vagyunk. Ha pedig netán maga Attila lenne ez a személy, akkor mindannyian Attila sarjai vagyunk. Furcsán hangzik, és látszólag szembemegy mindazzal, amit eddig írtam? Pedig valójában nagyon is logikus.
Bolyongás a családfaerdőben
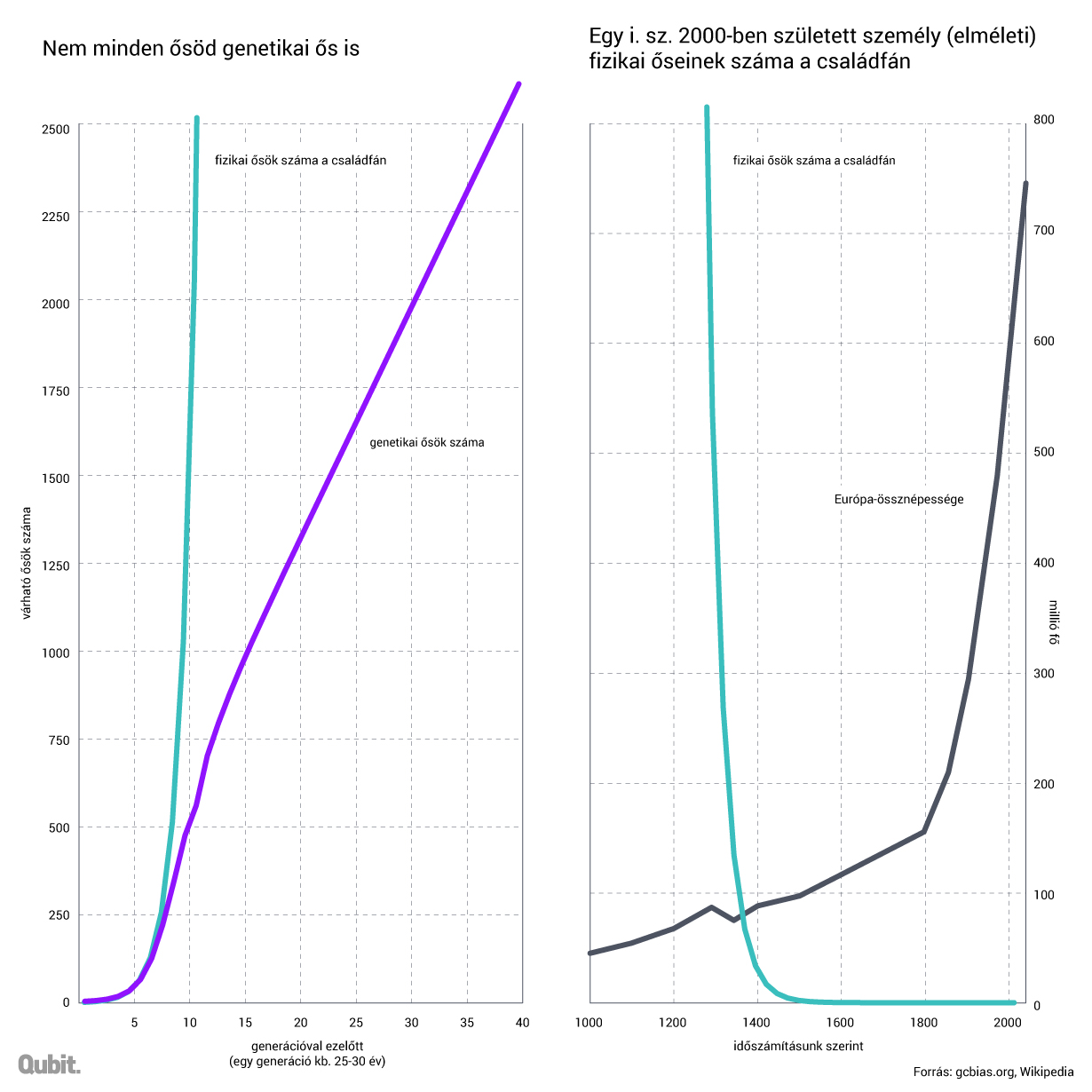
Az viszonylag könnyen belátható, hogy ugyan két szülőnk, négy nagyszülőnk és nyolc dédnagyszülőnk van, ám ez a fajta tisztán exponenciális növekedés nem folytatható. Ha így lenne, akkor tíz generációval ezelőtt 2048 felmenőnk lett volna, ezer évvel ezelőtt pedig milliárdnyi ősig vezetne vissza családfánk, miközben ekkor Európa teljes lakossága alig haladta meg a félmilliót, és a Földön sem élt még milliárdnyi ember. Ez azt jelenti, hogy a családfa sok ponton önmagába kell visszahajoljon, vagyis bizonyos személyek mind az apai, mind az anyai ágon őseink. Nem kell megrettenni, ha nem annyira extrém módon történik mindez, mint az egyiptomi fáraóknál vagy a Habsburg dinasztia spanyol ágánál, akkor nincs a dolognak komolyabb (genetikai) következménye sem.
Minél távolabb megyünk vissza az időben, annál több ilyen többszörös közös őst lelünk, mígnem durván ezer évvel ezelőtt kvázi Európa összes lakóját ott találni (egész pontosan azt a nagyjából 80 százalékot, akinek becsléseink szerint ma is élnek utódjai). Ráadásul nem csak a saját családfánk esetében, de minden más európainál is. Ez azt jelenti, hogy nem csak Attilának, de ha nem szakadt meg a leszármazási vonaluk, akkor Széphajú Haraldnak, Nagy Theuderichnek, Ptolemaiosznak és jó eséllyel a Rushdie-regénybeli ibn Rusdnak vagy épp Decebalnak is a közös utódai vagyunk. Az igencsak szapora Nagy Károlynak például biztosan, ugyanis ebben az esetben legalább egy, a közelmúltban elhunyt személyről tudunk, aki egészen a német-római császárig tudta a családfáját anyai ágon visszavezetni: ez pedig nem más, mint a köztudatba a Gyűrük Ura Szarumánjaként, illetve a Csillagok Háborúja Dooku grófjaként bekerülő Christopher Lee.7 De a napokban III. Béla feltételezett maradványainak vizsgálatairól publikált eredmények alapján jó eséllyel az Árpád-háznak is vannak férfi ágon ma élő leszármazottai.8 Ha pedig ez megerősítést nyer, azt is jelenti, hogy mindannyian (mindenki a kontinensen) leszármazottai vagyunk Árpádnak is. Persze ettől még nem beszélhetünk az ezer évvel ezelőtti Európa összes lakójáról genetikai ősünkként. Többségüktől biztosan nem örököltünk egyetlen bázispárnyi DNS-t sem, ettől ugyanakkor persze teljes értékű őseink.
A kontraintuitív összefüggést először bizonyító Joseph Chang és csoportja kimutatta, hogy ha egy generáció alatt egy-egy város lakosainak úgy 5 százaléka vándorol el, illetve egy-egy ország lakosainak 0,05 százaléka, az már elegendő ahhoz, hogy az összes ma élő ember családfája egy közös személybe konvergáljon nagyjából időszámításunk előtt 1415 tájékán.9 E modell értelmében nincs szükség olyan ugrásokra, hogy mondjuk valaki a koraközépkori Szombathelyről Alexandriába vándoroljon: a szomszédos területek közti, sok generáción át megvalósuló alacsony fokú, de folytonos vándorlás is elegendő ahhoz, hogy végül mégiscsak megjelenjenek a közös ősök a családfákon. Ha pedig léteznek nagyobb távolságokat áthidaló, majd ott sikeresen utódokat nemző egyének vagy csoportok (és a történelem tele van ilyen példákkal, még ha ez az utódnemzés gyakran konszenzus nélkül történt is, hogy eufemisztikusan fogalmazzunk), tovább gyorsítja a folyamatot.
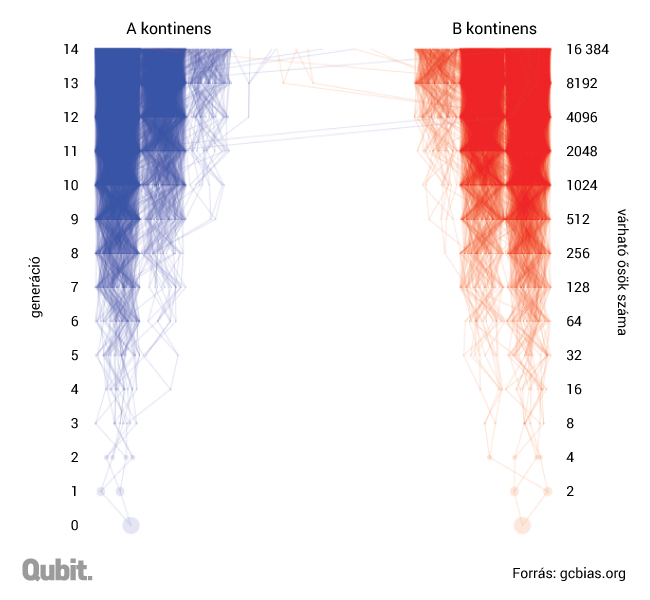
Lehetne ez egy elegáns, de hibás matematikai absztrakció is, hiszen a biológiai rendszerek nem feltétlenül követik pontosan a matematikai modellek logikáját. A University of California Graham Coop vezette kutatócsoportja azonban pár évvel ezelőtt, 2257 európai személy genomadatait felhasználva igazolta Changék modelljének helyességét.
Kódexmásolás és a DNS-blokkok
Az eredmények elemzése előtt még térjünk vissza az előző cikkünkben kifejtett hasonlathoz, hogy lássuk, miképpen zajlik az a bizonyos keveredés a homológokkal rendelkező kromoszómák anyaga között. (Emlékeztetőül: azt feltételeztük, hogy valamikor a 15. században egy spanyol kolostorban az Európában később csak Averroesként ismert andalúz filozófus ibn Rusd eredeti szövegét másoló írnok hibát ejtett, de ez nem tette érthetetlenné a szöveget, így ezt később sem veszi senki észre. A hibás másolatról készült másolatok elkerülnek olasz, francia és német kolostorokba is, ahol eltérő helyeken más és más hibákat vétenek a szerzetesek.)
Most annyival bonyolítjuk a korábban vázolt történetet, hogy nem egyetlen, hanem egy 22 kötetes mű sorsát követjük nyomon. Tegyük fel, hogy egy dunántúli kolostorban szert tettek az Averroes-mű két példányára is: a 22 kötetes opusz egyik verziója francia és német kolostorok kitérőjén keresztül érkezett, a másik pedig közép- és észak-olasz tudásközpontok érintésével. A korábban már részletezett okok miatt ezek nem pont egyforma verziók, kisebb-nagyobb különbségek fedezhetők fel bennük az írnokok pontatlansága miatt. Amikor egy morvaországi kolostorba kell továbbküldeni a művet, valami egészen furcsa dolog történik: miután a 44 kötetet (vagyis az olasz és a francia-német verziót is) lemásolják (esetleg új hibákat is ejtenek bennük), többé-kevésbé véletlenszerűen a verziók teljes és részfejezeteit is kicserélik, sőt egyes lapokat is. A 22 pár vegyes kötetből végül páronkét egyet-egyet küldenek tovább. Az úgynevezett meiózis jelensége annyival bonyolultabb, hogy nem csak a bizonyos dunántúli kolostorban készítik elő ilyen furán a továbbküldendő köteteket, hanem mindenütt így járnak el: egyre kisebb egységekre hullanak szét az olasz és német-francia földről érkező művek fejezetei, keverednek a más helyekről érkező példányokkal, végül azok a hibák, amiket mondjuk Észak-Olaszországban vétettek, bizonyos kiadásokból ki is vesznek.
A DNS-blokkok analógiájaként szolgáló teljes és részfejezetek, illetve oldalak eredete feltárható: a spanyol-német-francia(-magyar) vagy spanyol-olasz(-magyar) út során összeszedett írnoki hibák alapján. Ugyanígy a kromoszomális DNS-ben is felfedezhetünk olyan apróbb változatokat, polimorfizmusokat, amelyek együtt öröklődnek. És ha két külön személy genomját vizsgálva találunk ilyen közös mutációkat hordozó egységet (DNS-blokkot), akkor abban is biztosak lehetünk, hogy nekik közös genetikai ősük is volt. Minél közelebb élt ez a személy múltban (vagy minél több ilyen közös ős van a két családfán), annál több ilyen közös blokk jelenhet meg a két genomban.
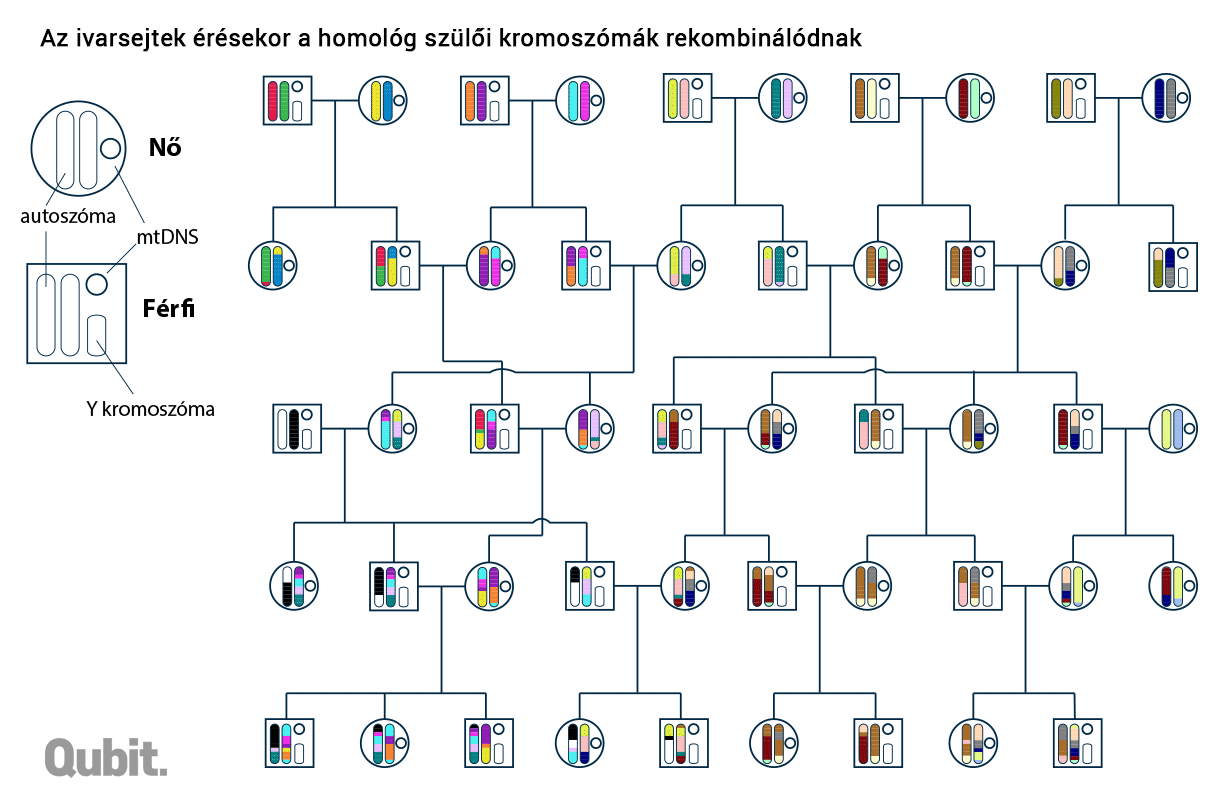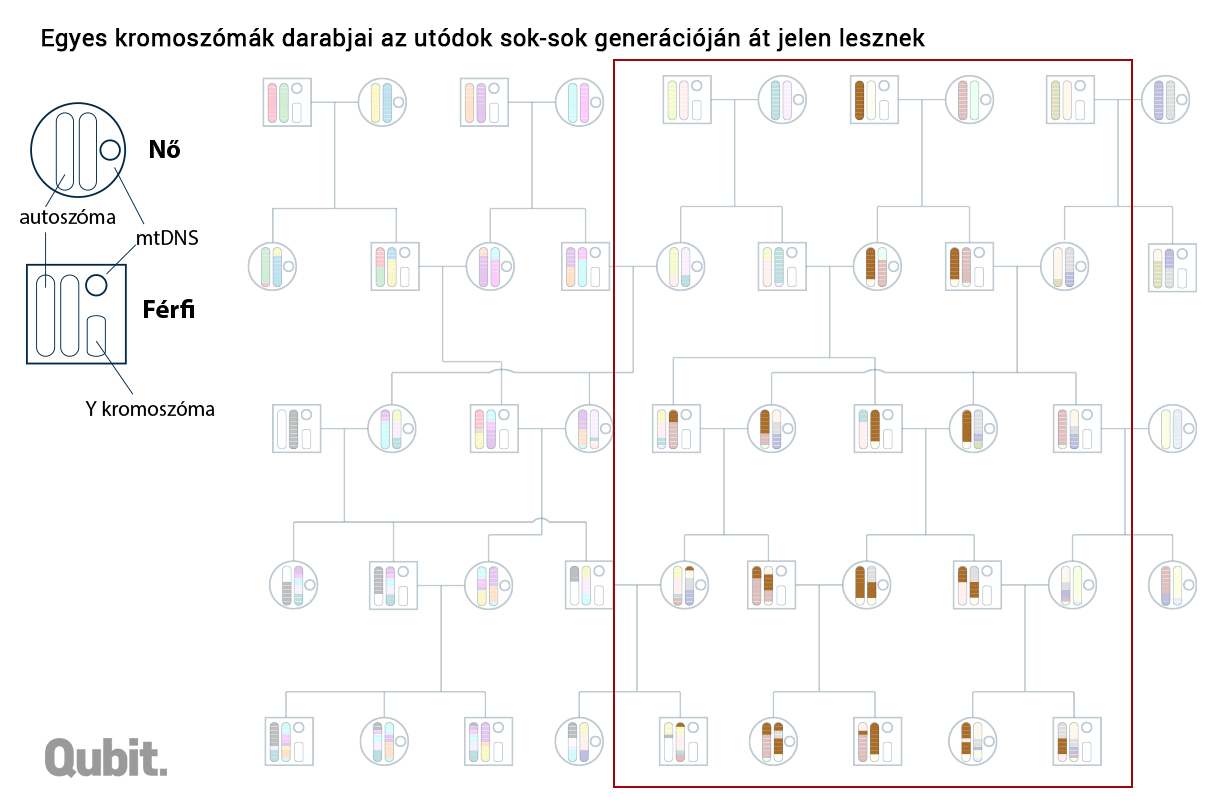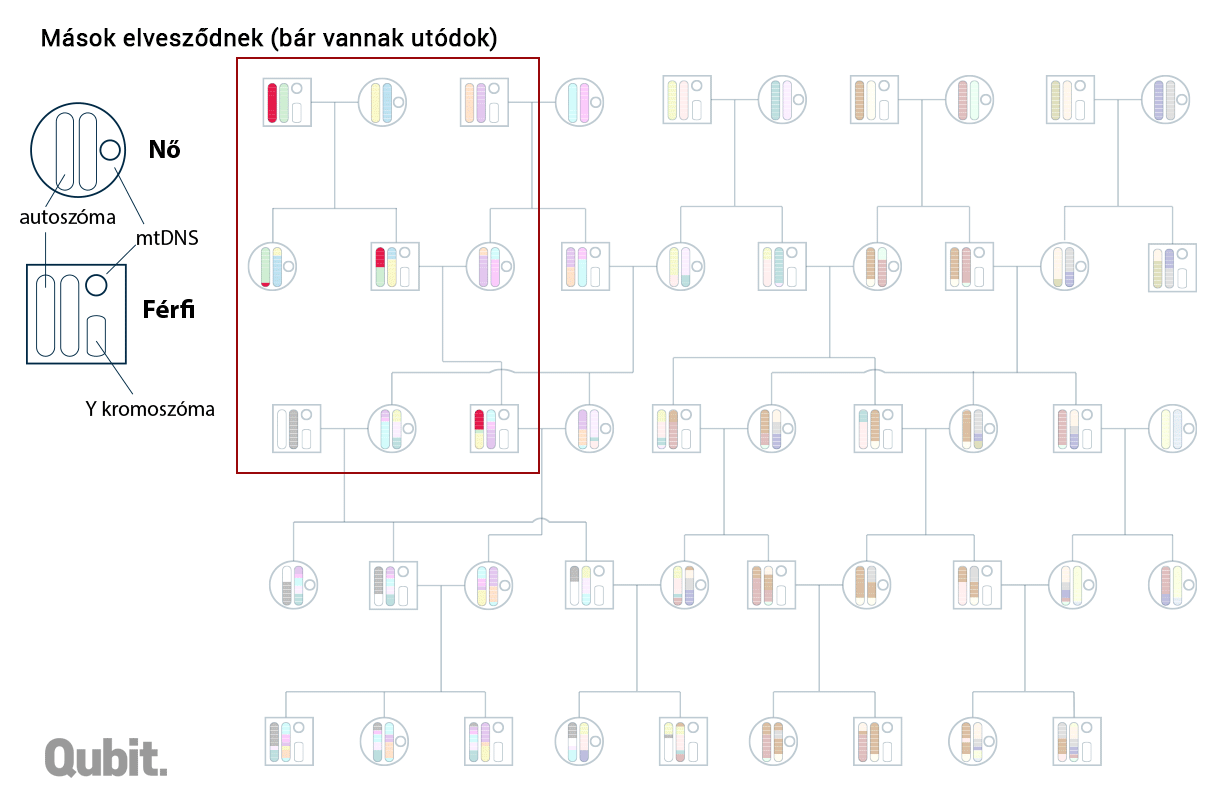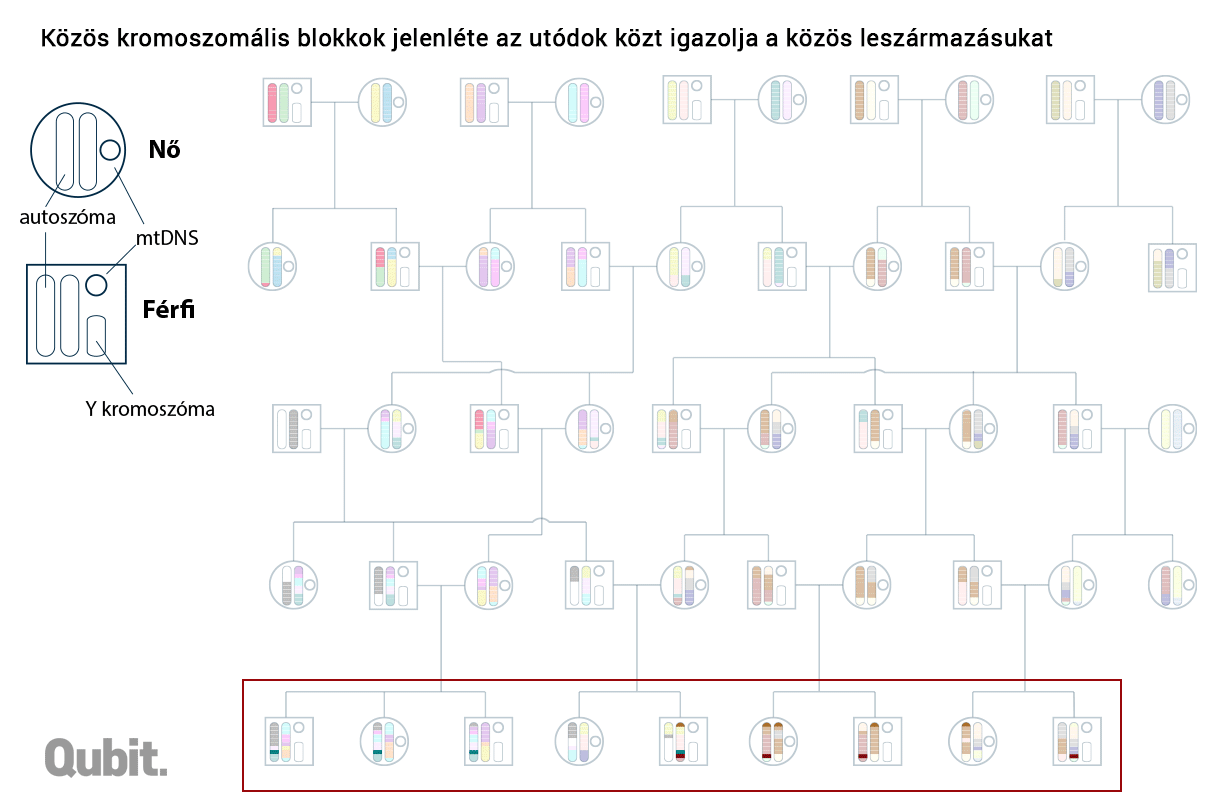
Közeli rokonság és távoli hasonlóság
Graham Coop és csoportja ezeknek a DNS-daraboknak a vizsgálata során fedezte fel, hogy az európai kontinens egész távoli sarkaiban (például az Egyesült Királyságban és Törökországban) élő személyek is osztoznak közös genetikai szegmenseken – az esetek bő 20 százalékában. Vagyis Changék matematikai modellje elég pontosan megfelel a valóságnak.10
Ha ugyanazok voltak az őseink, akkor miért nem vagyunk genetikailag szinte teljesen azonosak? Sejthető a válasz: mert a korabeli őseink közül csak kevesen őseink genetikailag is. Egy mai izlandi és egy ciprusi genetikai őseinek halmaza már csak alig fedi egymást. A távolabbi helyeken született ősök eleve kevesebben vannak, és bár a folyamatos migráció miatt a genetikai anyaguk folyamatosan bekeveredik a mi szűkebb közösségünk genetikai állományába, az örökítő anyag kíméletlen és folyamatos rekombinációja miatt nagyobb az esélye, hogy végül ez elvesszen, pontosabban nagyon kicsi az esélye (bár ezért nem lehetetlen), hogy annyira elterjedjen, hogy egy későbbi generáció többsége hordozza azt. A fizikai közelség miatt gyakoribb az is, hogy a kiterjedtebb családfák kaotikus kuszaságában már nem számon tartott negyed-, ötöd-, nyolcad-unokatestvérek egymással kötnek házasságot, így növelve az esélyét, hogy a valamikori közös ősök egyben a helyben élők genetikai ősei is legyenek, a tőlük származó DNS-blokkok nagyobb eséllyel fennmaradjanak.11

Bár az identitásunkat nem határozza meg, jó tudni, hogy mindannyian Nagy Károly és Kleopátra, a vikingek és az avarok, a hunok és a dákok, királyok és jobbágyok, zseniális polihisztorok és kétbalkezes bűnözők utódai vagyunk, vagyis szinte mindenki az ősünk, aki ezer évvel ezelőtt Európában, illetve a kétezer évvel ezelőtti ókori világ kevésbé eldugott szegletében élt. Mai identitásunkra nincs hatással, ha egy bonyolult és szerteágazó fa egyetlen távoli pontját azonosítjuk, mint ahogy többnyire az sem, ha esetleg valakitől konkrétan öröklünk pár tízezer bázispárnyi, legnagyobb valószínűséggel kódoló funkcióval nem is rendelkező DNS-blokkot. Így lettek a hunokból gepidák, a gepidákból avarok, és az avarokból magyarok, osztrákok, szerbek vagy épp románok, miközben térségünk genetikai összetétele már csak lassan, kisebb léptékben változott. Ezek a kisléptékű változások azonban izgalmas és fordulatos történelmi eseményekről, valamint genetikai véletlenekről tanúskodnak, amelyek feltárása a következő évtizedek archeogenetikájának lesz a feladata.
(A szöveg szakmai lektora Szécsényi-Nagy Anna, nyelvi lektora pedig Víg Julianna volt. Külön köszönet munkájukért.)
Lábjegyzetek:
[1] Az újkőkori keveredés témáját bővebben is feldolgoztuk egy korábbi impakták epizódban:
[2] Az emberi genom egyik legteljesebb molekuláris elemzését végző ENCODE projekt 2012-ben közzétett eredményei szerint az emberi genom több mint 80 százaléka valamilyen „funkciót” lát el (Maher 2012). A sokat vitatott eredmények részben szemantikai vitát eredményeztek: mint nem sokkal később kiderült, az ENCODE kutatói a „funkció” definicióját igencsak lazára vették, és tulajdonképpen minden molekuláris háttérzajnak is funkciót tulajdonítottak. Vagyis például az olyan helyeket is „funkcionálisnak” tekintették, ahol gyengén és következmények nélkül, de kapcsolódott egy transzkripciós faktor a genomhoz (mert a DNS szekvenciája emlékezetett a valódi kötőhelyére), miközben aligha lehet azt mondani, hogy ezeknek a helyeknek valódi feladata lenne a genom működésében. Azóta többfajta konzervatív becslés is született arra vonatkozóan, hogy mekkora része lehet valóban fontos az emberi genomnak, annak alapján, hogy a fontos szekvenciák felett őrködő szelekció mekkora részét védi a genomnak (kb. 8,2 százalék) (Rands et al. 2014), vagy hogy a jelenlegi mutációs gyakoriság és utódszám mellett mekkora rész lehet elméletileg szelekció alatt (kb. 10-15 százalék) (Graur 2017). Ezek egyértelmű következtetése, hogy az emberi genom nagyon nagy része nem rendelkezik konkrét feladattal a működésben. A vitáról bővebben: Doolittle and Brunet 2017.
[3] Az egyszerűség kedvéért nem foglalkozunk itt a genetikai betegségek nagyon tragikus és egyébként igen fontos kategóriájával, amelyek egyébként gyakran (de nem mindig) valóban monogénes formában öröklődnek.
[4] A testmagasság genetikájáról: (Lello et al. 2017).
[5] A fülcimpaalak genetikájának részletes vizsgálata: (Shaffer et al. 2017).
[6] Az utalás Illyés Gyula Ki a magyar? zárógondolatára vonatkozik (Illyés 1982).
[7] Christohper Lee színész és Nagy Károly rokonságáról a University College Dublin díjátadóján is megemlékeztek.
[8] A III. Béla maradványaiból kivont DNS-t elemző tanulmány (Olasz et al. 2018).
[9] Joseph Chang szimulációi (Rohde, Olson, and Chang 2004).
[10] Graham Coop csoportjának releváns cikke a közös DNS-blokkok öröklődéséről: (Ralph and Coop 2013).
[11] A témáról még rengeteget lehet írni és beszélni, és ha valakit jobban is érdekelnek ezek a kérdések, akkor a Graham Coop honlapján található GYIK remek kiindulás az elmélyüléshez.
Referenciák:
Doolittle, W. Ford, W. Ford Doolittle, and Tyler D. P. Brunet. 2017. “On Causal Roles and Selected Effects: Our Genome Is Mostly Junk.” BMC Biology 15 (1). https://doi.org/10.1186/s12915-017-0460-9.
Graur, Dan. 2017. “An Upper Limit on the Functional Fraction of the Human Genome.” Genome Biology and Evolution 9 (7):1880–85.
Illyés Gyula. 1982. Ki a magyar.
Lello, Louis, Steven G. Avery, Laurent Tellier, Ana Vazquez, Gustavo de los Campos, and Stephen D. H. Hsu. 2017. “Accurate Genomic Prediction Of Human Height.” https://doi.org/10.1101/190124.
Maher, Brendan. 2012. “ENCODE: The Human Encyclopaedia.” Nature 489 (7414):46–48.
Olasz, Judit, Verena Seidenberg, Susanne Hummel, Zoltán Szentirmay, György Szabados, Béla Melegh, and Miklós Kásler. 2018. “DNA Profiling of Hungarian King Béla III and Other Skeletal Remains Originating from the Royal Basilica of Székesfehérvár.” Archaeological and Anthropological Sciences. https://doi.org/10.1007/s12520-018-0609-7.
Ralph, Peter, and Graham Coop. 2013. “The Geography of Recent Genetic Ancestry across Europe.” PLoS Biology 11 (5):e1001555.
Rands, Chris M., Stephen Meader, Chris P. Ponting, and Gerton Lunter. 2014. “8.2% of the Human Genome Is Constrained: Variation in Rates of Turnover across Functional Element Classes in the Human Lineage.” PLoS Genetics 10 (7):e1004525.
Rohde, Douglas L. T., Steve Olson, and Joseph T. Chang. 2004. “Modelling the Recent Common Ancestry of All Living Humans.” Nature 431 (7008):562–66.
Shaffer, John R., Jinxi Li, Myoung Keun Lee, Jasmien Roosenboom, Ekaterina Orlova, Kaustabh Adhikari, 23andMe Research Team, et al. 2017. “Multiethnic GWAS Reveals Polygenic Architecture of Earlobe Attachment.” American Journal of Human Genetics 101 (6):913–24.

