Titokzatos mikroorganizmusok, az archeák jelölik ki a földi élet legvégső határait

Az 1980-as évek óta ismert a kutatók számára, hogy egyes mikroorganizmusok a legextrémebb földi viszonyok között is képesek túlélni. A mélytengeri hidrotermális kürtőket, hőforrásokat, extrém kémhatású vagy rendkívül sós környezeteket úgynevezett extremofil – azaz szélsőséges környezeteket eltűrni képes vagy egyenesen azokat kedvelő – mikrobák lakják.
A Nature Ecology & Evolution tudományos folyóiratban tavaly novemberben megjelent tanulmány arra keresett válaszokat, hogy léteznek-e olyan mikrobák, amelyek képesek megélni egyszerre erősen savas, nagy sókoncentrációjú és forró hőforrásokban. A francia és spanyol kutatókból álló csoport Jodie Belilla vezetésével arra is választ szeretett volna kapni, hogy az extrém környezeti viszonyok közötti túléléshez szükséges evolúciós adaptációk inkompatibilisek-e egymással - ami megmagyarázná, hogy miért nem sikerült meggyőző módon eddig savas, erősen sós hőforrásokban bármilyen élőlényt találni.

A szakemberek kérdéseik megválaszolásához kutatásaikat az etiópiai Afar törésvonalon található Dallol hidrotermális területen és környékén végezték, ahol erős és gyorsan változó intenzitású hőforrások találhatók. Ezek nagy sókoncentrációjú (33–50 százalékos), magas hőmérsékletű (25–110 Celsius-fokos) és erősen savas (alacsony kémhatású) tavakat, tavacskákat hoznak létre. A Dallol teljesen földön kívülinek tűnő környezete így elég jó analógiája például a NASA Spirit marsjárója által 2007-ben vizsgált egykori hidrotermális terület viszonyainak.
Az ehhez hasonló kutatások tehát arra is fényt deríthetnek, hogy milyen korlátok között képes alkalmazkodni az élet az extrém környezetekhez a Földön és más bolygókon.
Extrém hő, sómennyiség és pH – egyszerre sok
A szakemberek mintáikat három expedíció során, 2016-ban, 2017-ben és 2018-ban gyűjtötték - a Dallol-területen belül négy különböző zónából. A víz- és talajmintákon aztán környezeti DNS-vizsgálatokat, mikrobiális tenyésztést, áramlási sejtszétválasztást (FACS) és pásztázó elektronmiszkroszkópos (SEM), valamint kémiai vizsgálatokat alkalmaztak.

Az első mintavételi zóna a Dallol-dóm tetején lévő színes tavacskákban volt, amelyeknek a vize erősen sós (37–42 százalék), rendkívül savas (pH 0–1) és helyenként akár 108 Celsius-fokos hőmérsékletű. A második zóna közeli, nagy sókoncentrációjú kanyonokból állt, ahol a kutatók barlangi sós tavacskákból és rövid ideig létező pocsolyákból vettek mintákat. A Dallol-dóm alján található állóvizek nagy sókoncentrációjúak (35 százalék), közepes hőmérsékletűek (30 Celsius-fok) és kissé savasak voltak (pH 4-6). Ezzel ellentétben a nagyon sós, forró és savas „Fekete-tóhoz” közeli pocsolyák nem túl meglepő módon savasabbak (pH 3), melegebbek (40 Celsius-fok) és sósabbak (35–60 százalék) voltak.
A harmadik zóna a „Sárga-tavat” és környékének pocsolyáit foglalta magában. Ezek extrém savasak, (pH 1,8), melegek (40 Celsius-fok) és erősen sósak (50 százalék fölötti sókoncentráció) voltak. Az utolsó, negyedik mintavételi zóna pedig az erősen sós (36 százalék), szinte semleges kémhatású (pH 6,5) és alacsonyabb hőmérsékletű Assale-tavat foglalta magában, amelyet az előbbi három zónával való kevésbé extrém összehasonlítási alapnak használtak a szakemberek.
A minták közt három alapvetően eltérő kémiai környezetet azonosítottak. Az első zóna tavacskái a kémiai vizsgálatok alapján konyhasó, azaz nátrium-klorid (NaCl) által túlszaturált sóoldatok, nagy mértékű vastartalommal. A második és negyedik zónában szintén NaCl-tartalmú pocsolyákat találtak, de ezekben kevesebb vasat mértek a kutatók. A második és harmadik zóna „Fekete-tóhoz” és „Sárga-tóhoz” közeli részeinél a tavacskák nagyon magas magnézium- és kalciumkoncentrációt mutattak. Ez utóbbi területeken a sejtmembránroncsoló anyagok koncentrációja olyan magas a szakemberek mérései szerint, hogy azok eleve kizárják bennük bármilyen élet jelenlétét.
Ahhoz, hogy megállapítsák, hogy található-e élet az adott mintavételi helyeken, és ha igen, milyen rendszertani csoporthoz tartozik, genetikai vizsgálatokat alkalmaztak. A Dallol-dóm tavacskáiból, valamint a Fekete- és Sárga-tóból vett DNS-minták klasszikus, laboratóriumi baktériumszennyeződéseket mutattak, további baktériumokat pedig a kutatók szerint a területet bejáró turisták vagy a szél hurcolhatták a tavacskákba.
A legjobb törekvéseik ellenére a kutatóknak csak a második zóna barlangi tavaiból, valamint a kevésbé extrém környezetet biztosító Assale-tóból sikerült valóban ottani létformákból származó DNS-t izolálniuk. A molekuláris biológiában általánosan használt polimeráz-láncreakció (PCR) segítségével megsokszoroztak meghatározott DNS-szakaszokat a mintákban, rengeteg másolatot készítve róluk. A megsokszorozott DNS-szakaszok a fehérjeszintézisért felelős molekuláris gépezetnek, azaz a prokarióta riboszómának a 16S és 18S riboszomális RNS-komponenseit kódoló génjei voltak. Ezen gének használata régóta bevett gyakorlat mikrobák azonosítására, rendszertani kategorizálására vagy evolúciós kapcsolataik feltárására (ami a filogenetika tárgykörébe tartozik).
A genetikai vizsgálatok eredményeiből kiderült, hogy a barlangi tavacskák és az Assale-tó területén rendkívül változatos archeák élnek.
De mik azok az archeák?
Ahhoz, hogy megértsük, milyen mikrobákat találtak ezeken a területeken a kutatók, nagyobb kitérőt kell tennünk.
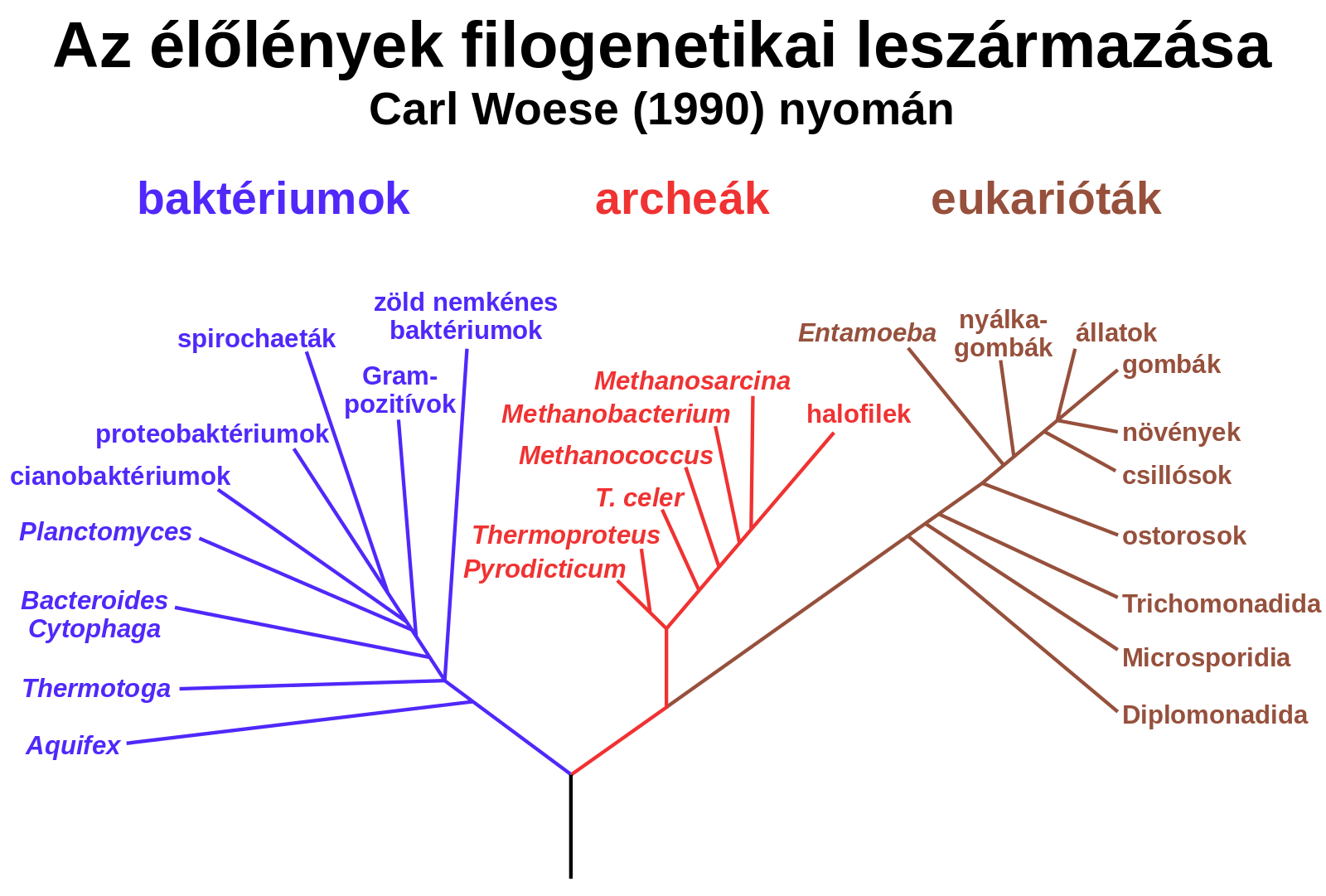
A földi élővilág három nagy csoportra, úgynevezett doménre osztható: baktériumokra, archeákra és eukariótákra. A baktériumokat kórokozó szerepük miatt nem kell senkinek bemutatni. Ez az egyébként nagyon változatos csoport viszonylag egyszerű felépítésű, sejtmag nélküli, azaz prokarióta sejtekkel rendelkezik, és több mint 3,5 milliárd éve jelen van bolygónkon. Az eukarióták komplex, sejtmaggal rendelkező sejtből vagy sejtekből állnak, és közel kétmilliárd évvel ezelőtt jelentek meg a Földön. Az eukarióták egy archea gazdasejt és egy baktérium fúziójából jöttek létre egy szimbiogenezisnek nevezett folyamat során. Az egykori baktériumból alakult ki később a mitokondrium, az eukarióta sejtek kulcsfontosságú sejtszervecskéje, amely ATP (adenozin-trifoszfát) formájában kémiai energiát állít elő. Az eukarióták közé tartozunk mi is, ahogy minden állat, növény, gomba, illetve rengeteg egysejtű életforma is.
Az archeák a baktériumokhoz hasonlóan nagyon ősi, sejtmag nélküli mikroorganizmusok, felületesen nézve nem sokban különböznek tőlük. Molekuláris, sejttani eltéréseik, illetve evolúciós történetük alapján viszont egyértelműen megkülönböztethetők, és a földi élet története szempontjából az egyik legfontosabb csoportot alkotják. Erre Carl R. Woese és kollégái ébredtek rá 1977-ben, amikor publikálták a genetikai vizsgálatokon alapuló törzsfájukat, bevezetve ezzel az előbb ismertetett három doménes felosztást. Míg az archeák sejtmembránja teljesen egyedi az élővilágban, a fontos genetikai rendszereiket működtető fehérjéik az eukariótákkal állnak közeli rokonságban.
Bár először extrém élőhelyeken, például felszíni vagy mélytengeri hőforrásokban azonosították őket, valójában mindenhol jelen vannak, még az emberi szervezetben is. Kései felfedezésük egyik fő oka, hogy lényegében egyetlen biztosan betegséget okozó – azaz patogén – archea sem ismert. A legismertebbek közülük talán az Euryarchaeota csoportba tartozó metanogén archeák, amelyek anyagcseréjük melléktermékeként metánt állítanak elő.
A Belilla és kollégái által azonosított archeák fele a Haloarchaea (korábbi nevükön Halobacteria) csoportba, negyedük pedig a Nanohaloarchaeák közé tartozik. Mindkettő a nagyobb Euryarchaeota csoport tagja, életmódjukat tekintve pedig sókedvelő extremofilnek tekinthetők. A mintákban azonosított fennmaradó mikrobák szinte a teljes archea-változatosságot lefedik. A szakemberek szerint a nagy mennyiségű Haloarcheán Nanohaloarcheák élnek úgynevezett ekto-szimbionta kapcsolatban, egyfajta külső élősködőként.
A 2010-es évek közepének evolúciós genetikai eredményei elkezdték árnyalni az élővilág korábban bemutatott háromosztatúságának képét, így ma azt mondhatjuk, hogy a földi élet két elsődleges csoportból, baktériumokból és archeákból áll.
Az eukarióták, tehát mi magunk is az archeák közé tartozunk.
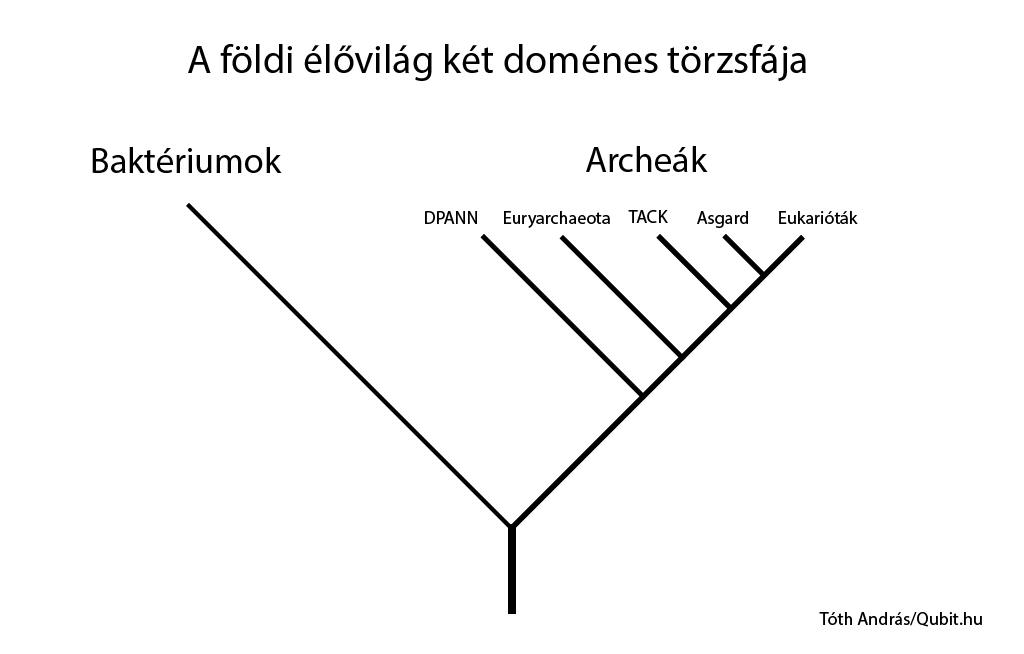
Érdemes ugyanakkor hozzátenni, hogy az eukarióták létrejötte forradalmi esemény volt a földi élet történetében. Az eukarióták az archea őseikben szépen lassan létrejött genetikai és sejtbeli innovációkat felturbózták, majd elképesztően kiterjesztették - köszönhetően egyebek mellett a bekebelezett baktérium (későbbi mitokondrium) által szolgáltatott energiatöbbletnek. Ez az eseménysor – amelynek pontos lefolyását a mai napig heves viták övezik – tette lehetővé a komplex élőlények és végül saját fajunk kialakulását a Földön.
Módszertani kérdőjelek
Belilla és kollégáinak tenyésztéses vizsgálatai, amelyekkel laboratóriumi körülmények közt szerették volna szaporodásra bírni a mintákban lévő életformákat, a genetikai vizsgálatokkal egybevágó eredményt hoztak: a Dallol-dóm, a Sárga- és a Fekete-tavak területéről származó mintákban semmilyen aktivitást nem tapasztaltak. Ugyanerre jutottak a DNS-t kereső fluoreszcencia-mikroszkópos és pásztázó elektronmikroszkópos (SEM) vizsgálatok. Mikrobákat csak a korábban említett barlangi tavacskákból és a közelükben lévő sós síkságokról származó mintákban sikerült észlelni és tenyészteni.
A Nature Ecology & Evolution folyóiratban a tanulmánnyal együtt egy John E. Hallsworth mikrobiológus által írt kritika is megjelent. Ebben Hallsworth kifejti, hogy Belilláék eredményei az élettelen sós-savas vizek vonatkozásában egybevágnak a korábbi, extrém sókoncentrációjú vizekben végzett kutatásokkal.
Hallsworth azonban három fontos észrevételt is megfogalmazott a tanulmánnyal kapcsolatban, amelyek az ilyen kutatásokban általánosan felhasznált módszerek korlátait is mutatják:
- Az összegyűjtött DNS-minták csak rendszertani információt hordoztak, és nem lehet belőlük megállapítani, hogy a sejtek valóban aktívak voltak-e a mintavétel idején.
- A mintavétellel nyert oldatminták víztartalmának mérése 25 Celsius-fokon, és nem a mintavételi forrás helyi (in situ) hőmérsékletén történt.
- A mintákban lévő sejtmembránroncsoló oldott anyagok valamint az oldatok kémhatásának mérése pontatlan a felhasznált mérési módszer miatt.
Ennek ellenére Hallsworth kiemeli, hogy Belilla és kollégái sikeresen leírt nagyszámú, erősen sós tavacska geokémiáját és mikrobiális gazdagságát – amely a kémiai tulajdonságok széles skáláját is mutatja. Ezzel pedig Hallsworth szerint a kutatócsoport feltárta a jelenlévő archea mikrobaközösség változatosságát.
Emellett a kutató szerint a tanulmány további fontos kérdéseket is felvet arra vonatkozóan, hogy képesek-e a mikrobák fenntartani anyagcseréjüket, ha az élőhelyükként szolgáló sós oldatok víztartalma alacsonyabb, mint az az osztódásukhoz szükséges. Illetve egyáltalán élőhelynek tekinthető-e egy környezet, ha elméletileg élő sejtek vannak benne, de azok nem képesek lényegi anyagcserét végezni? Az extremofil adaptációkkal kapcsolatban is felmerül, hogy ha egyes sók is több stresszfaktort képviselnek egy mikroba számára, akkor definíció szerint egy sótűrő mikrobának több extrém környezeti tényezőhöz kell képesnek lennie alkalmazkodni?
Óvatosabbnak kell lenni a földön kívüli élet keresésekor
Belilláék munkájának azonban van még egy fontos konklúziója, amely óvatosságra int mind az ősi földi mikrobafosszíliák, mind az egykori földön kívüli élet keresésekor (például a Marson).

A kutatók a legextrémebb helyekről vett minták áramlási sejtszétválasztásos (FACS) vizsgálatakor is sejtekre emlékeztető képződményeket találtak. Elektronmikroszkóppal és röntgen-spektrométerrel aztán alaposabb vizsgálatoknak vetették alá ezeket a gömb és henger alakú struktúrákat, és megállapították, hogy többnyire szilíciumkristályokból, vas-alumínium szilikátokból és más ásványokból álló biomorfokról, azaz élettelen (abiotikus) eredetű, sejtekre hasonlító képződményekről van szó.
A kutatók munkájuk eredményéből három fontos konklúziót emelnek ki:
- Két fő kémiai korlátját azonosították az élet fennmaradásának folyékony víz jelenlétében: magas magnéziumion koncentrációjú sóoldatokat, valamint egyszerre nagyon savas és magas sókoncentrációjú oldatokat. A szerzők szerint ez arra utalhat, hogy az extrém kémhatású és sókoncentrációjú környezetben való túléléshez szükséges genetikai adaptációk egymással inkompatibilisek.
- Bár az extrém környezetek többnyire nem mutatnak nagy változatosságot az ott élő életformák tekintetében, a kutatók itt éppen az ellenkezőjét tapasztalták. Ahol volt élet, ott nagyon különböző csoportokhoz tartozó archeákat találtak. Ez arra utal, hogy ezek mind rendelkeznek az extrém sós környezethez nélkülözhetetlen adaptációkkal, amelyek legalább részben egymástól függetlenül jöhettek létre az evolúció során.
- A nem biológiai eredetű, főleg szilíciumban gazdag sejtekre emlékeztető formációk gyakorisága óvatosságra int a mikrofosszíliák értelmezésével kapcsolatban, és szükségessé teszi, hogy a vizsgálatok több független módszerrel erősítsék meg az egykori élet jelenlétét, akár a korai Földről, akár a Naprendszer más bolygóin zajló kutatásokról van szó.
Ez a kutatás tehát közelebb visz minket annak megértéséhez, hogy milyen környezeteket képes még a földi élet tolerálni, és miket nem. Az evolúció nagyon más irányokat vehet a különböző bolygókon, mégis joggal feltételezhetjük, hogy kémiai felépítésüket nézve nagy vonalakban a földi szénalapú életre emlékeztető létformák lehetnek jelen a Földön kívül. Mindez kritikusan fontos információ, amikor a Marson, a külső Naprendszerben lévő holdakon vagy annál is távolabb vizsgálják az adott környezet jelenlegi vagy múltbeli élhetőségét.
Belilla, J., Moreira, D., Jardillier, L. et al. Hyperdiverse archaea near life limits at the polyextreme geothermal Dallol area. Nat Ecol Evol 3, 1552–1561 (2019) doi:10.1038/s41559-019-1005-0
Hallsworth, J.E. Microbial unknowns at the saline limits for life. Nat Ecol Evol 3, 1503–1504 (2019) doi:10.1038/s41559-019-1021-0
Kapcsolódó cikkek a Qubiten:

Így találhatnánk legkönnyebben földön kívüli életet a Naprendszerben
A Szaturnusz holdjának gejzírjeiből való mintavétel páratlan lehetőséget kínál annak eldöntésére, hogy létezik-e élet az Enceladus jégkérge alatti óceánban.

Nyúl vagy kacsa? Hogyan értelmezzük a hiányzó ősmaradványokat?
Vajon mennyire képesek a fosszíliák megbízható információkat nyújtani a földi élet történetének pontos alakulásáról és az evolúció mechanizmusairól? Attól függ, honnan nézzük, és melyik tudóst kérdezzük, írja Bárdos Dániel, akinek kutatási területe a paleontológia filozófiája.

Magyar kutatók is hozzájárultak a felfedezéshez, miszerint a Marson egykor lehetett élet
A jövőre induló Mars 2020 misszió landolási helyszíne egy 3,6 milliárd évvel ezelőtt létező folyó helye lesz, ahol a marsi élet szempontjából kulcsfontosságú üledékeket keres majd a NASA marsjárója.


#/media/File:The_hydrothermal_system_of_Dallol,_Ethiopia.png){kind=link}