A földi élet megismételhetetlen ugrása: az eukarióták megjelenése

A földi életet hatalmas szakadék választja ketté. Ez nem túl feltűnő, mivel a szakadék messze a szemünk felbontóképessége alatt, az egysejtűek világában húzódik. Minden élő, amit magam körül látok, algától jómagamig, sejtmagvas (eukarióta) sejtekből áll. Az iskolai tankönyvek jobbára néhány keresetlen sorral intézik el a jóval egyszerűbb, sejtmag nélküli egysejtűeket, a baktériumokat és archaeákat. Ez persze érthető, hiszen csak az eukarióta sejteknek sikerült olyan csodálatosan összetett lényeket létrehozniuk, amelyek akár a bolygót is el tudják hagyni, ahol kialakultak. Azóta, hogy esélyünk van másutt is életet találni a Naprendszerben, a tudományt különösen érdekelni kezdte, hogy vajon találhatunk-e fejlett életet odakint, illetve, hogy idehaza, a Földön miért csak egyetlen egyszer alakult ki a magasabb rendű, eukarióta élet. Hogyan sikerült a baktériumoknak átugrani a szakadékon? A tudomány legújabb eredményei szerint egyáltalán nem biztos, hogy ehhez az ugráshoz hatalmas energiatöbbletre volt szükség, ahogy azt eddig hittük. Még az is elképzelhető, hogy az átmenet nem villámgyors ugrás, az akadály pedig nem is szakadék volt.
Áthidalhatatlan szakadék?
A ma élő élőlények két csoportja között hatalmas komplexitásbeli különbség van. Csak a sejtmagvasok (azaz eukarióták) értek el magasabb rendű szerveződést, mint a komplex többsejtűség vagy az egyik sejt másikba költözése (endoszimbiózis). A sejtmag nélküliek (baktériumok és archaeák, együtt a prokarióták), néhány kivételtől eltekintve nem képesek bonyolultabb szerveződésre, gyakorlatilag mind egysejtűek. Amióta ez a különbség nyilvánvalóvá vált, azóta érdekli az embereket, hogy vajon mikor, hogyan és főleg miért ugrottak át a prokarióták ezen a szakadékon. Az első kettő kérdésre elég jó válaszaink vannak, és eddig azt hittük, az utolsóra is. Egy új tanulmány azonban, amely a Nature Ecology & Evolution folyóiratban jelent meg 2022-ben, fontos adalékkal szolgál ahhoz, hogy megkérdőjelezzük a miértre adott korábbi válaszokat. Úgy tűnik, az eukarióta eredet oka keményebb dió, mint gondoltuk.
A prokarióta és eukarióta szervezetek közötti szakadék a bakteriológia hajnala óta izgatja a kutatókat. Különösképpen azóta, hogy Roger Stanier és Cornelius van Niel mikrobiológusok a prokariótákat és az eukariótákat strukturálisan alapvetően különböző, de evolúciósan folytonos csoportokként értelmezték. Korábban ez utóbbi sem volt egyértelmű, de a szaporodó bizonyítékokra alapozva a kutatók úgy gondolták, hogy a két csoport közül az egyik ősibb volt (kézenfekvően az egyszerűbb prokarióták), és belőlük fejlődött ki evolúcióval a másik csoport, az eukarióták. Azóta számos bizonyíték igazolta, hogy a baktériumok nagyon korán kialakultak a Földön, és az eukarióták később, folytonos leszármazással jöttek létre belőlük. Azonban az igazán érdekes az, hogy úgy tűnik, a prokarióták a földi életük 4 milliárd éve alatt összesen egyszer voltak képesek a szakadékot átlépni. Egyre pontosabb filogenetikai adatok alapján tudjuk, hogy ez kb. másfél-kétmilliárd éve történt. De vajon mi tartotta vissza őket ennyi ideig? És miért csak egyszer jött el az eukarióta megváltás? Miért nem volt azóta második, harmadik eukarióta advent?
Sajnos az egysejtűek korai evolúciós eseményeivel az a baj, hogy nem fosszilizálódnak túl jól. Az eukarióták hajnalát sűrű köd fedi, és nem ismerjük az átmeneti formákat, így a köztes lépések sorrendjét sem. Ezért kapnak nagyon nagy szerepet az elméleti, modellező kutatások. A korábbi hipotézis szerint a prokariótákat az egyszerű felépítésükből fakadó energetikai plafon tartja vissza a komplexitásnövekedéstől, és ahhoz, hogy ezt áttörjék, elengedhetetlen szükségük volt a mitokondriumra, ami közkeletű nézet szerint az eukarióta újítások energetikai ‘adu ásza’ („the powerhouse of the cell”). Az említett 2022-es tanulmány szerzői, Paul Schavemaker és Sergio Muñoz-Gómez azonban arra a következtetésre jutottak, hogy a mitokondrium nem feltétlenül volt elengedhetetlen az eukariótává váláshoz, főleg, ha figyelembe vesszük, hogy kezdetben aligha lehetett olyan hatékony, mint a mai „erőművek”. De akkor mi tette lehetővé az eukariótáknak a szakadék feletti átjutást és a rákövetkező megismételhetetlen sikert?
Ahhoz, hogy ezt jobban megértsük, előbb meg kell értenünk, hogy miféle strukturális és energetikai előnye van annak, ha valaki eukarióta király a prokarióta pórnép közt.
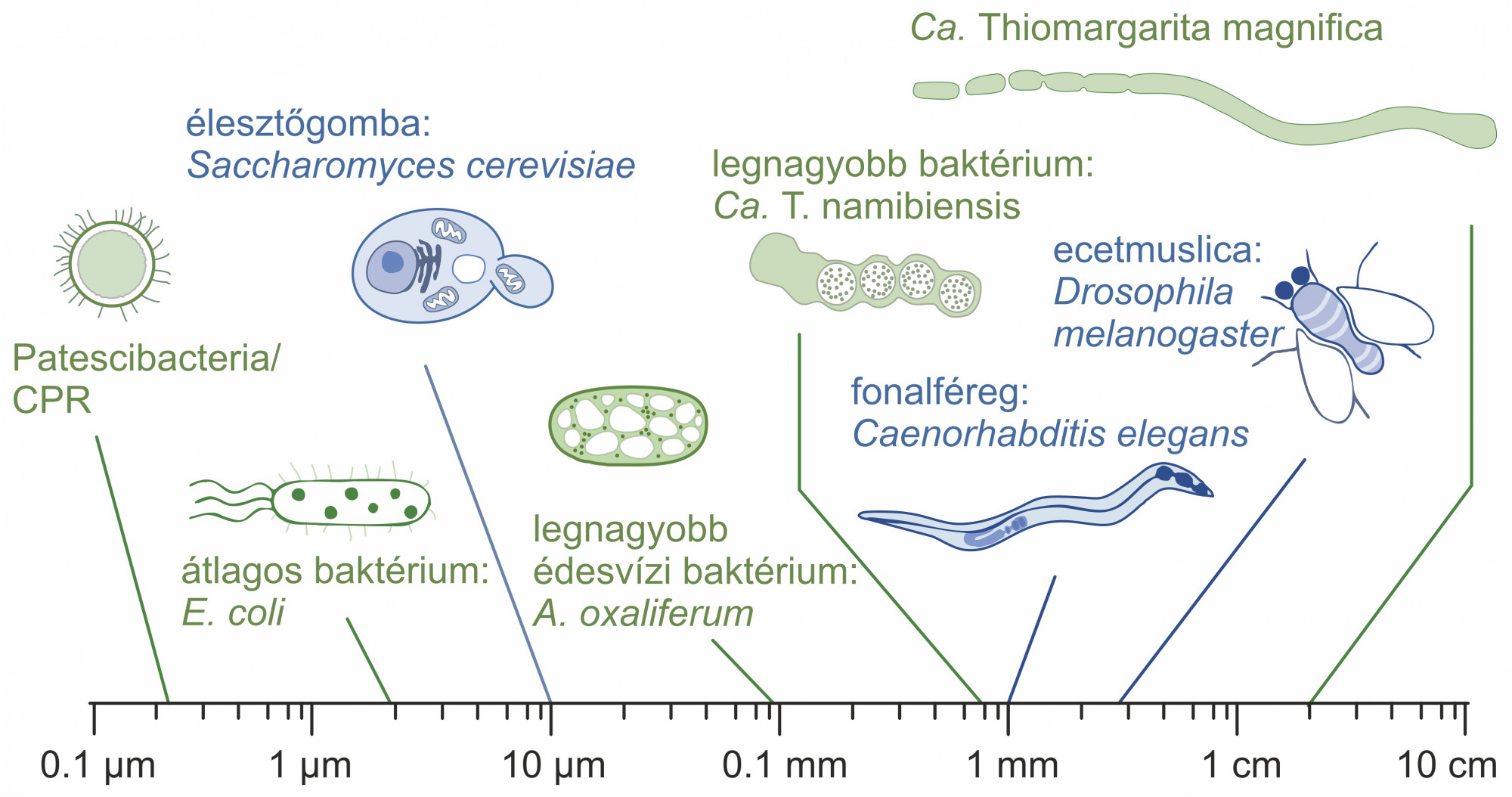
Az energetikai plafon a szakadék mögött
Az eukariótává válás egyszeri volta és az átmenetek hiánya azt sugallják, hogy az ilyesmi ritka, de gyors lefolyású esemény lehet. A ritkaság oka valamilyen rejtett faktor, amely megakadályozta (és most is akadályozza) a további, független eseményeket, ahol a prokarióták átlépik a komplexitásküszöböt. Az eukariótává válás általános vélekedés szerint nagyon szorosan összefüggött a mitokondrium megszerzésével. Érdekes módon az is egyedüli esemény volt, akár a sejtmag és az eukarióták eredete. Miért nincs prokarióták közötti gyakrabban a mitokondriumhoz hasonló endoszimbiózis? Egy friss (még nem elbírált) tanulmány egy régi hipotézist cáfolva arra a következtetésre jutott, hogy a ritkaság oka nem lehet a partnerek anyagcsere-összeférhetetlensége: gyakorlatilag bármely két prokariótát kiválasztva azok (elméletben) képesek lehetnek egymást segítve szaporodni. Nick Lane és Bill Martin szerint a szakadék az eltérő sejtfelépítések eredménye, amelyek miatt meghatározó különbség van az eukarióták és a prokarióták energiatermelésében.
Dióhéjban a különbség lényege az energiatermelő (bioenergetikai) membránok méretében rejlik. Minden önellátó prokarióta sejt a teljes sejtfelszínén keresztül folytat energiatermelést, legyen szó akár fotoszintézisről, akár sejtlégzésről. És minden prokarióta sejt egyetlen kromoszómában tárolja a teljes örökítőanyagát, azaz egy genomhoz a sejtfelszínnel arányos energiatermelés jár. Ezzel szemben az eukarióta sejtek akár több tízezer mitokondriummal is rendelkeznek, amelyek mindegyike önállóan termeli az energiát a saját, erősen megnövelt belső membránfelületén. Mindeközben az eukarióta sejteknek ugyanúgy egyetlen, központi genomjuk van, bár az kétségkívül nagyobb, mint a prokariótáké. Ezért az eukarióta sejt energiatermelése nem a felszínnel, hanem durván a térfogattal arányos, ami – ahogy a sejt növekedik – köbösen nő, miközben a felszín csak négyzetesen. De ez még nem lenne elég különbség, hiszen e szerint ugyanakkora térfogatban sok prokarióta sejt ugyanannyi energiát tudna megtermelni, mint egy nagy eukarióta sejt (lásd az alábbi ábrát).
A modern mitokondriumok elképesztően költséghatékonyak. Bár részben autonómok, nem kell a teljes bakteriális struktúrájuk fenntartását megfizetniük: ugyan van saját DNS-ük, de csak a legszükségesebb fehérjéket gyártják helyben, a többit a gazdasejt termeli nekik. Képesek önállóan energiát termelni, miközben a sejtmag gyakorol központi kontrollt felettük (azaz például nem szaporodhatnak kényükre-kedvükre). Tulajdonképpen úgy működnek, mint valami kihelyezett fiókintézmény, amelyet az anyagyár tart fenn. A mitokondriumok így tekintélyes költséget takarítanak meg, pontosan úgy, mint a fiókvállalatok. És a mitokondriumok bárhol lehetnek a sejtben, nincsenek a sejthártyához kötve. Magyarán azonos sejtméretben egy eukarióta sejt több energiát tud megtermelni, mint egy prokarióta.
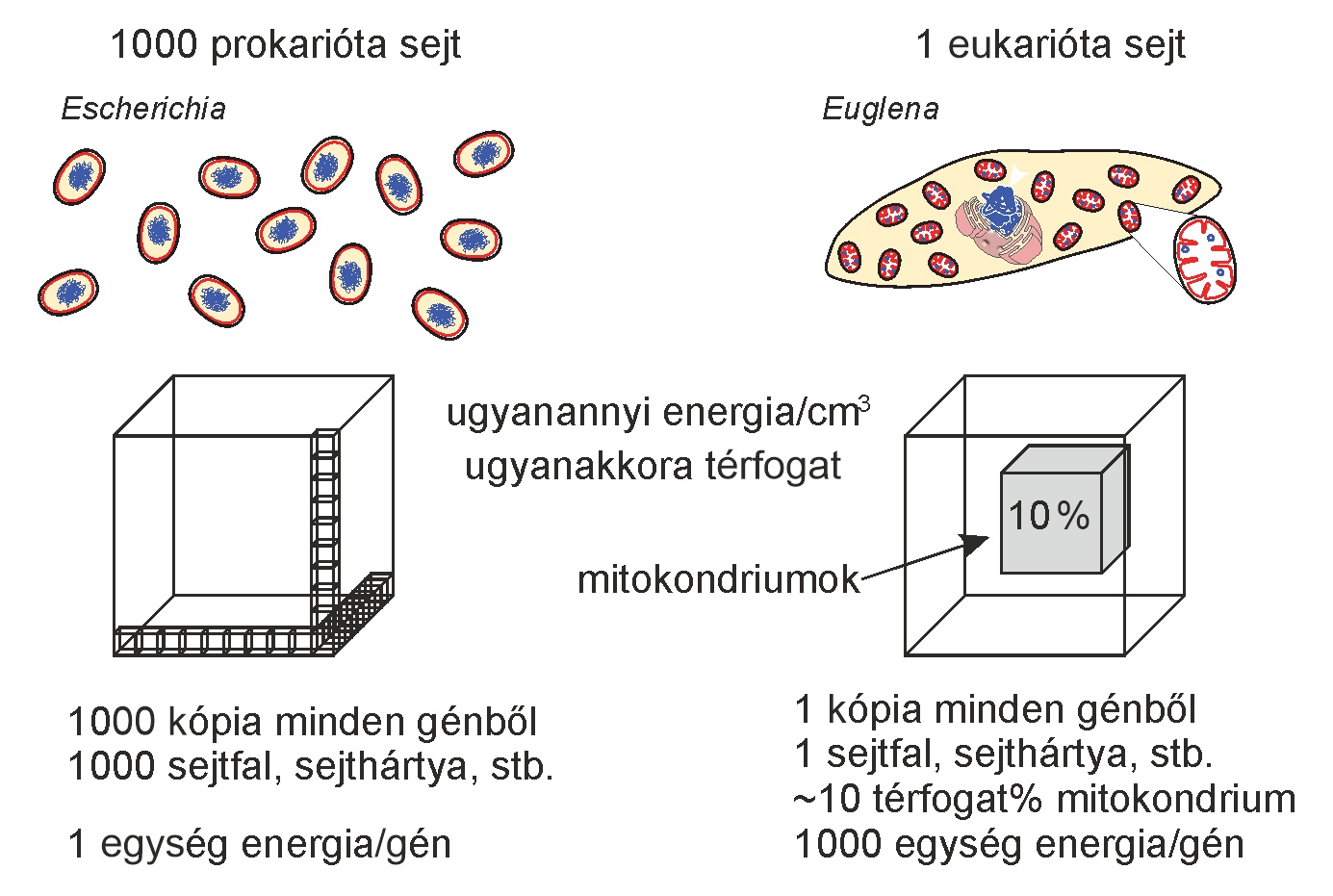
A modern eukarióta felépítés tehát előnyös a prokariótákéhoz képest, akiknek az energiatermelése membránkötött és így nem a térfogattal, hanem a sejtmembránnal skálázódik. Ők is növelhetik a sejtméretüket, de azonos térfogatban kevesebb energiát tudnának termelni, mint egy mitokondriumokkal telipakolt eukarióta. Ráadásul egy bizonyos méreten felül már nem lehet gyorsan és hatékonyan eljuttatni a termelt fehérjéket a membránhoz a nagy távolság miatt. Itt játszik szerepet a mitokondriumok félautonómiája, no meg a fejlett eukarióta sejtváz. E logisztikai probléma megoldására a prokariótáknak számos genommásolatot kellene kihelyezniük a sejthártya mentén, hogy lokálisan el tudják látni a teljes bioenergetikai membrán fehérjeigényét. Ez rendkívül költséges, éppen ezért nem is látjuk gyakran.
Vannak azonban kivételek. Egy újonnan leírt, óriás Thiomargarita baktérium pontosan ezt teszi: centiméter hosszú sejtjei vannak, és a membránja mentén több mint félmillió genetikailag aktív sejtszervecske található (tulajdonképpen kihelyezett génközpontok), amelyek valószínűleg helyben felelnek a sejtlégzés fehérjéinek termeléséért. A Thiomargarita kiválóan példázza, hogy egy prokarióta sejt nem növekedhet akármekkorára, mert a költségei a térfogattal skálázódva sokkal gyorsabban nőnek, mint a sejtfelszínen megtermelt energia. Ezzel szemben a mitokondriumok lehetővé tették eukarióta gazdáiknak, hogy azok nagyobbra nőhessenek. Azonban korántsem egyértelmű, hogy pontosan mikortól nyújtották ezt az előnyt.
Lane és Martin szerint ez a strukturális különbség nem csak a prokarióták és eukarióták közötti komplexitásbeli szakadékért felelős. Úgy vélik, hogy a modern eukarióták számára elérhető génenkénti (genomonkénti) energia százszor vagy akár ezerszer is nagyobb, mint a prokarióták esetében. De ennél is tovább mennek: szerintük a szakadékot csakis egy olyan endoszimbionta megszerzésével lehetett áthidalni, amely képes volt ezt a többletet biztosítani. Vagyis a mitokondrium nem csak oka a jelenleg megfigyelhető szakadéknak, hanem egyúttal azért is felelős, hogy egy ősi prokarióta egyszer átugorhatta ezt a szakadékot – az, amelyik megszerezte a mitokondriumot. Vagyis szerintük a prokarióta komplexitást egy energetikai plafon korlátozza, az eukariótává váláshoz ezt át kellett törni, és ehhez nélkülözhetetlen volt a mitokondrium által nyújtott többletenergia. Ezt nevezték el energetikai hipotézisnek. De vajon tényleg nélkülözhetetlen energiaforrás volt a mitokondrium?
A komplexitásbeli szakadék, illetve a prokarióta fejlődést korlátozó energetikai plafon megfigyelhető tény. Igazolja ezt a baktériumok kis mérete és a kivételes felépítésű óriásbaktériumok is, mint a Thiomargarita. Ez azonban még nem feltétlenül jelenti, hogy valóban energetikai akadály gátolja a prokariótákat abban, hogy eukarióta szintű komplexitást érjenek el, mondjuk kisebb méretben. Még kevésbé jelenti azt, hogy a szakadék áthatolhatatlan lenne mitokondriumok nélkül. A mitokondrium és a sejtmag látszólagos egyidejű megjelenése félrevezető lehet: az egyidejűség nem feltétlenül jelent evolúciós okozatot. Valójában az összes fő eukarióta vívmány (mitokondriumok, sejtmag, aktív sejtváz, fagocitózis stb.) ugyanabban a homályos, ködlepte időszakban jött létre, amiről semmilyen sorrendiségre utaló egyértelmű nyom nem maradt fenn.
Létrával minden plafon elérhető
Többen meg is kérdőjelezték Lane és Martin hipotézisét az abszolút energetikai akadályról. Elvégre léteznek hatalmas baktériumok, és találtak olyan egysejtű eukariótát is, ami kiválóan megél mitokondrium nélkül. Határozottan úgy tűnik, a két domén részben átfed a rendelkezésre álló energia és komplexitás tekintetében (lásd a 3. ábrát). Az energetikai hipotézis ráadásul nem ad magyarázatot arra, hogy ha ekkora sikert jelentett egy légzésre képes baktérium bekebelezése, akkor miért nem történik ilyesmi gyakrabban a prokarióták világában?
Részletesebb energetikai vizsgálatok elnagyoltnak találták Lane és Martin számításait, melyet meglehetősen kevés eukarióta (12) és prokarióta (55) példára alapoztak. Több részletesebb tanulmány is arra a következtetésre jutott, hogy az energetikai akadály a modern prokarióták és eukarióták között nem létezik, vagy legalábbis nem olyan hangsúlyos. Michael Lynch és Georgi Marinov 2017-es tanulmányukban egyenesen azt állították, hogy nincs okunk azt feltételezni, hogy a membrán bioenergetikája közvetlenül szerepet játszott volna a prokarióták eukariótává válásában. Schavemaker és MuñozGómez most megjelent tanulmányukban is pontosan azt vizsgálták meg, hogy vajon ténylegesen mennyi többletenergia nyerhető egy korai eukarióta számára, aki már bekebelezte a mitokondrium ősét, de még nem olvasztotta be annak génjeit a saját genomjába.
A kutatópáros izgalmas új elemzése teszteli Lane és Martin energetikai hipotézisét, elsősorban a szimbionta membrán méretére és a genetikai felépítésére összpontosítva. A kutatók kimutatták, hogy a modern eukarióták genetikai felépítése (egyetlen gazdagenom, több redukált szimbionta genom) valóban előnyös a modern prokariótákhoz képest (akiknek több genommásolatot kell fenntartaniuk, ahogy a sejt mérete növekszik). Ezek az eredmények megerősítik a feltételezést, hogy az eukarióták energiatermelése előnyösebben skálázódik a mérettel, mint a tisztán prokarióta megoldások, amelyek szigorúan a membrántól függenek.
Azonban eredményeik szerint a modern eukarióta egysejtűek előnye messze nem akkora, ahogy azt Lane és Martin állította. A tanulmány szerint az eukarióták energetikai többlete mindössze kb. háromszorosa a modern prokariótákénak, szemben a Lane és Martin által javasolt, igen tekintélyes 200 000-szeres előnnyel. Persze már egy háromszoros relatív különbség is jelenthetett abszolút előnyt a korabeli egysejtűek között. De figyelem, a kritikus kérdés nem az, hogy mekkora előny érhető el mai mitokondriumokkal, hanem hogy mekkora előnyt jelentettek az ősi, kevésbé hatékony formáik – főleg, ha nem is voltak. Ne feledjük, hogy egyelőre az sem biztos, hogy az első eukariótáknak volt mitokondriuma.
És itt durran igazán nagyot Schavemaker és MuñozGómez matematikai modellje. A kutatók megvizsgálták, hogy mennyi többletet hozhatott a konyhára egy olyan korai partner, amely még nem volt olyan „költséghatékony erőmű”, mint a mai mitokondriumok, azaz nem kizárólag az energiatermelésre specializálódott, és nem olvadt be genetikailag a gazdagenomba. Az új számítások szerint a korai szimbiontának tulajdonított energianyereség teljesen elpárolgott. Vagyis a korabeli prokarióták és proto-eukarióták között nem volt áthidalhatatlan energetikai szakadék, és a korai mitokondrium-hordozó sejteket ugyanúgy korlátozta az energetikai plafon, mint a prokariótákat. A mitokondrium és az eukarióta szerveződés előnye csak jóval később érvényesülhetett – amikor a mitokondriális gének bekerültek a gazda genomba. Amint a mitokondriumok elvesztették genomjuk nagyobbik részét, már nem kellett a rengeteg kópia fenntartásának a költségét is fedezni (lásd az alábbi ábrát).
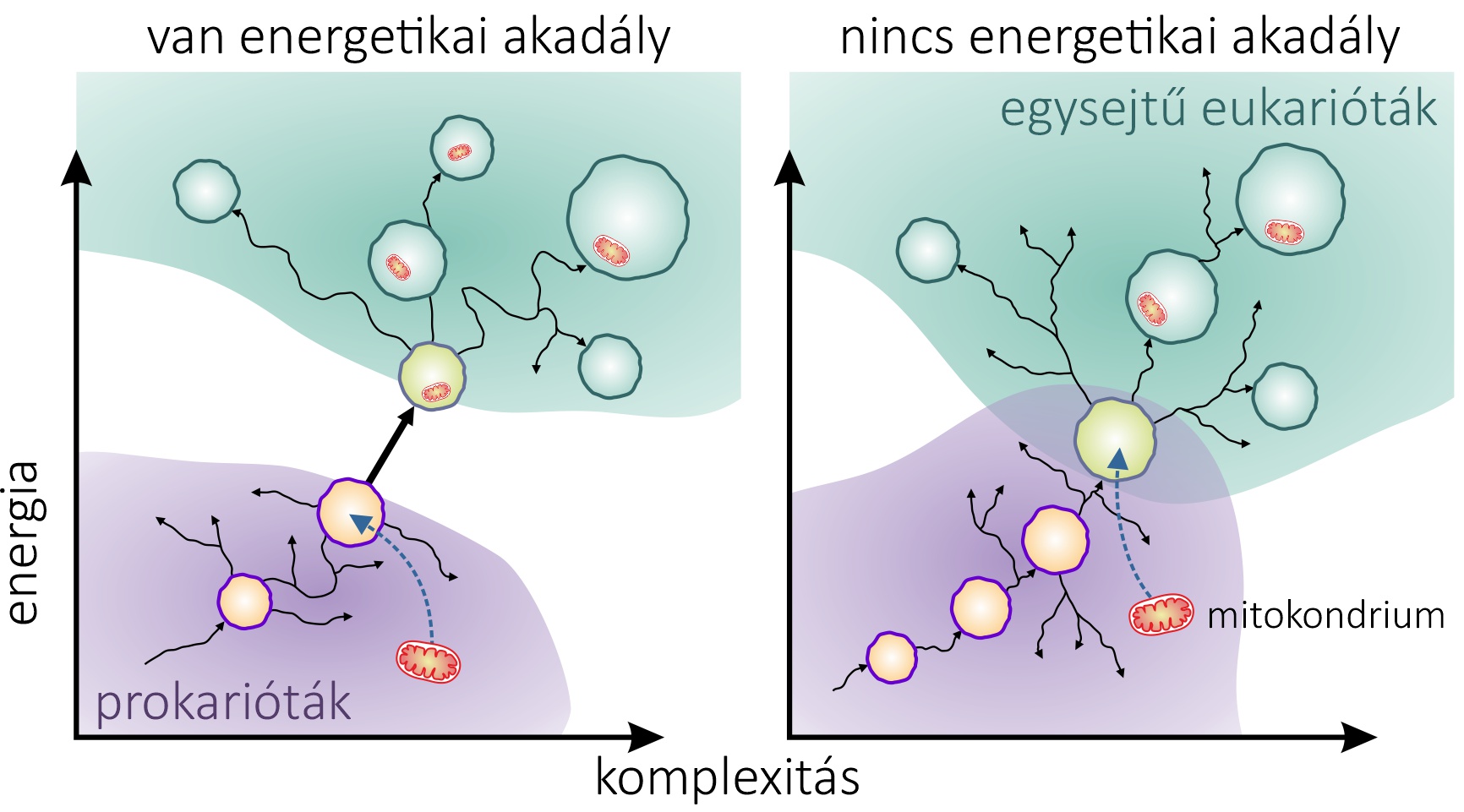
Mitokondrium először, másodszor, senki többet harmadszor
A páros eredményei tehát arra utalnak, hogy az energetikai szakadék a kapcsolat hajnalán nem volt jelentős – vagy talán nem is létezett. Ha pedig nem volt áthidalhatatlan szakadék, akkor vélhetően nem ez volt az eukariótává válás akadálya. Ez a konklúzió messzemenő következményekkel jár. Az energetikai korlát eltűnése a prokarióta és eukarióta domén között azt jelenti, hogy az archaea gazda fokozatosan fejlődhetett eukariótává, kisebb változásokkal szerezve előnyt a versenytársak felett. Ráadásul az átmenet nem feltétlenül támaszkodott a mitokondrium-nyújtotta korai előnyre, hanem fordítva: a megnövekedett komplexitás volt az, ami lehetővé tette a mitokondrium elfogását. Elképzelhető, hogy a mitokondrium azután jelent meg, hogy a korai eukariótákban már kialakult a sejtmag, a belső membránok vagy a fagocitózis (lásd az alábbi ábrát).
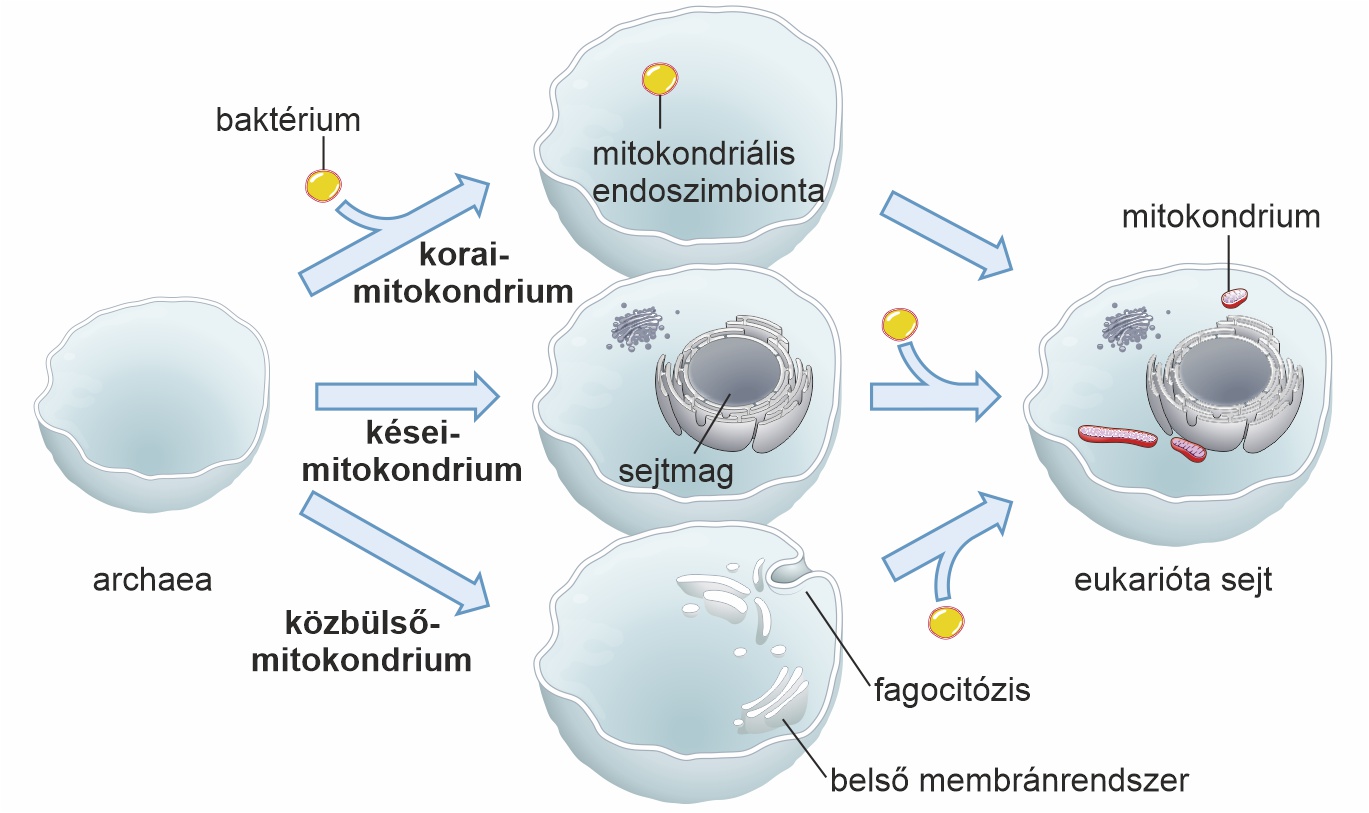
A prokarióták közt azonban a sejtmaghoz hasonló struktúra nem ismert. De a sejtmagvasok között sem találtunk olyat, aminek elsődlegesen hiányzik a mitokondriuma (azaz nem már létező szimbiontájukat vesztették el). Az átmeneti formák hiánya teret ad a spekulációknak. Sokáig azt gondoltuk, hogy a fagocitózis is olyan exkluzív eukarióta tulajdonság, mint a sejtmag, amelyet még csak megközelíteni sem tudtak a prokarióták, lévén nagyon költséges.
2019-ben azonban japán kutatók leírtak egy baktériumot, ami képes a fagocitáló eukariótákhoz hasonlóan (de tőlük evolúciósan függetlenül) táplálékszemcséket vagy akár kisebb sejteket is bekebelezni. A palaui mitológia éhes óriásáról elnevezett Uab baktérium bizonyítja, hogy kialakulhat és működhet bonyolult, kizárólag eukariótának hitt mechanizmus prokarióta hardveren is. A fagocitózis tökéletes korai megoldás lenne a mitokondrium eredetére, mivel enélkül nem világos, hogyan kerülhetett be a mitokondrium őse a gazdasejtbe.
De vajon miért volt szükség a mitokondriumra az elején, ha nem az energiatöbbletért? Ez a kérdés félrevezető. Helyesebb lenne azt kérdezni, hogy mi volt a mitokondrium kezdeti előnye, amikor bekebelezte a gazdasejt. Az energetikai hipotézis szerint úgy tűnik, mintha a gazda a jövőbeli előnyök érdekében fektetett volna be a mitokondriumba – a jövőbeli többletenergiáért vagy azért az evolúciós potenciáért, amit a többletenergia által nyújtott genetikai robbanás tett lehetővé. Azonban az evolúció nem invesztál a jövőbe; inkább a jelenlegi helyzetből törekszik kihozni a legtöbbet. A fagocitózisnak például van azonnali előnye: a ragadozó egysejtű megeheti a szomszédját, sőt, el is tárolhatja őket rosszabb időkre. Bármelyik hatalmas előnyt jelenthetett a versenytársakkal szemben.
Az eukarióta szingularitás
Talán a mitokondriumok nem voltak elengedhetetlenek a korai eukarióta tulajdonságok (beleértve a fagocitózis) kialakulásához, fenntartásához és fejlődéséhez. Lehet, hogy a gazda fagotróf volt, és így szerezte meg a mitokondriumot, ahogy az például az összes színtest esetében történt. Persze az is lehet, hogy a mitokondrium volt az első a sorban, és a korai eukarióták többször, egymástól függetlenül találták fel a fagocitózist. Ha így is volt, az az eredeti energetikai hipotézistől eltérő okok miatt történhetett, amit most már példák és számítások is megerősítenek.
Az is igaz ugyanakkor, hogy az eukarióták legközelebbi rokonai nem a baktériumok között keresendők (ahová az Uab is tartozik), hanem az archaeák, vagyis ősbaktériumok között, azoknak is az Asgard nevű csoportjában. Ezt a csoportot még nem annyira ismerjük, viszont rohamléptekkel zajlik a feltérképezésük, minden évben valamilyen új, érdekes eredménnyel. Nem is olyan régen sikerült csak először laborban szaporodásra bírni őket, amiből kiderült, hogy például a Prometeoarchaeum olyan szoros kapcsolatban él egy baktériumpartnerrel, hogy annak a hiányában nem képes növekedni. A korai mitokondriumot feltételező elméletek (az energetikai hipotézis is) pontosan azt feltételezik, hogy a gazdasejt a szimbionta ősével efféle szoros anyagcsere-kapcsolatban volt, például egyik a másik termékét fogyasztotta (szintrófia). Azonban ezek az elméletek nem tudnak számot adni arról, hogy az egyszerű gazda miképp kebelezte be a partnerét, fagocitózis nélkül.
A fagocitózishoz hasonló mechanizmus előfordulása prokariótákban (Uab), azt jelenti, hogy létezhetett kézenfekvőbb mód a korai partnerek fizikai egyesülésére annál, mint hogy két szintróf prokarióta spontán összeolvad. Ez utóbbira nem ismerünk példát, ami különösen annak fényében gyanús, hogy a szintróf kapcsolatok rendkívül gyakoriak a prokarióták világában. Sajnos mindkét út igen valószínűtlen és megfigyelhetően ritka jelölt a prokarióta endoszimbiózisok magyarázatára. Ezért aztán mindkét út tökéletesen alkalmas a kivételesen ritka (ne szépítsük, egyedülálló) eukarióta eredet magyarázatára. Azonban ahogyan csökken a prokarióták és eukarióták között feltételezett energetikai szakadék, úgy nyer egyre nagyobb támogatást a mitokondriumot megelőző, lépésről lépésre megszerzett archaea-bonyolultság forgatókönyve.
Az evolúció ritkán történik ugrásszerűen, legfeljebb mi érzékeljük hirtelennek a folytonos, apró változásokat a távolból nézve. Minél régebben történt az esemény, annál valószínűbb, hogy nem látjuk a köztes lépéseket. Carl Woese amerikai mikrobiológus már az 1970-es években amellett érvelt, hogy a most megfigyelhető szakadék látszólagos, és pusztán annak következménye, hogy mindkét leszármazási ágon kb. kétmilliárd évnyi független evolúció eredményét látjuk, azóta, hogy minden ma élő prokarióta és eukarióta őse elvált egymástól. A friss kutatások azt mutatják, hogy nem kell fejlett energiatermelő ős-mitokondriumot és azonnali jelentős energiatöbbletet feltételezni a kapcsolat elején ahhoz, hogy az első eukarióták evolúciós előnyt élvezzenek. Akár a szerényebb energetikai többlet, akár más, nem közvetlenül érvényesülő kezdeti előny (fagocitózis, farmolás) is elvezethetett hosszabb távon az eukarióták sikeréhez. Vak prokarióták között egy félszemű is lehetett király.
A szerző evolúcióbiológus, az Ökológiai Kutatóközpont Evolúciótudományi Intézetének főmunkatársa. Húsz éve kutatja a földi élet nagy átmeneteit, azon belül is az eukarióták eredetét. A cikk rövidebb formában eredetileg a Nature Ecology & Evolution lapban jelent meg: Zachar 2022 Closing the energetic gap, Nature Ecology & Evolution.
Hivatkozások
Betts, H. C., M. N. Puttick, J. W. Clark, T. A. Williams, P. C. J. Donoghue, and D. Pisani. 2018. Integrated genomic and fossil evidence illuminates life’s early evolution and eukaryote origin. Nature Ecology & Evolution 2:1556–1562.
Bremer, N., F. D. K. Tria, J. Skejo, S. G. Garg, and W. F. Martin. 2022. Ancestral state reconstructions trace mitochondria but not phagocytosis to the last eukaryotic common ancestor. Genome Biology and Evolution.
Cavalier-Smith, T. 2014. The neomuran revolution and phagotrophic origin of eukaryotes and cilia in the light of intracellular coevolution and a revised Tree of Life. Cold Spring Harbor Perspectives in Biology 6:1–31.
Chiyomaru, K., and K. Takemoto. 2020. Revisiting the hypothesis of an energetic barrier to genome complexity between eukaryotes and prokaryotes. Royal Society Open Science 7:191859.
Duve, C. de. 2007. The origin of eukaryotes: a reappraisal. Nature Reviews Genetics 8:395–403.
Ettema, T. J. G. 2016. Mitochondria in the second act. Nature 531:39–40.
Imachi, H., M. K. Nobu, N. Nakahara, Y. Morono, M. Ogawara, Y. Takaki, Y. Takano, K. Uematsu, T. Ikuta, M. Ito, Y. Matsui, M. Miyazaki, K. Murata, Y. Saito, S. Sakai, C. Song, E. Tasumi, Y. Yamanaka, T. Yamaguchi, Y. Kamagata, H. Tamaki, and K. Takai. 2020. Isolation of an archaeon at the prokaryote–eukaryote interface. Nature 577:519–525.
Jékely, G. 2007. Origin of phagotrophic eukaryotes as social cheaters in microbial biofilms. Biology Direct 2.
Karnkowska, A., V. Vacek, Z. Zubáčová, S. C. Treitli, R. Petrželková, L. Eme, L. Novák, V. Žárský, L. D. Barlow, E. K. Herman, P. Soukal, M. Hroudová, P. Doležal, C. W. Stairs, A. J. Roger, M. Eliáš, J. B. Dacks, Č. Vlček, and V. Hampl. 2016. A eukaryote without a mitochondrial organelle. Current Biology 26:1274–1284.
Lane, N., and W. F. Martin. 2010. The energetics of genome complexity. Nature 467:929–934.
Libby, E., C. Kempes, and J. Okie. 2022. Metabolic compatibility and the rarity of prokaryote endosymbioses. bioRxiv.
Lynch, M., and G. K. Marinov. 2015. The bioenergetic costs of a gene. Proceedings of the National Academy of Sciences 112:15690–15695.
Lynch, M., and G. K. Marinov. 2017. Membranes, energetics, and evolution across the prokaryote-eukaryote divide. eLife 6.
Martin, W. F. 2017. Symbiogenesis, gradualism, and mitochondrial energy in eukaryote origin. Periodicum Biologorum 119:141–158.
Martin, W. F., A. G. M. Tielens, M. Mentel, S. G. Garg, and S. B. Gould. 2017. The physiology of phagocytosis in the context of mitochondrial origin. Microbiology and Molecular Biology Reviews 81:8–17.
Schavemaker, P. E., and S. A. Muñoz-Gómez. 2022. The role of mitochondrial energetics in the origin and diversification of eukaryotes. Nature Ecology & Evolution.
Shiratori, T., S. Suzuki, Y. Kakizawa, and K. Ishida. 2019. Phagocytosis-like cell engulfment by a planctomycete bacterium. Nature Communications 10.
Stanier, R. Y. 1970. Some aspects of the biology of cells and their possible evolutionary significance. Pages 1–38 in H. P. Charles and B. C. J. G. Knight, editors. Organization and Control in Prokaryotic and Eukaryotic Cells, Twentieth Symposium of the Society for General Microbiology. . Cambridge University Press, Cambridge.
Stanier, R. Y., and C. B. van Niel. 1962. The concept of a bacterium. Archiv für Mikrobiologie 42:17–35.
Volland, J.-M., S. Gonzalez-Rizzo, O. Gros, T. Tyml, N. Ivanova, F. Schulz, D. Goudeau, N. H. Elisabeth, N. Nath, D. Udwary, R. R. Malmstrom, C. Guidi-Rontani, S. Bolte-Kluge, K. M. Davies, M. R. Jean, J.-L. Mansot, N. J. Mouncey, E. R. Angert, T. Woyke, and S. V. Date. 2022. A centimeter-long bacterium with DNA contained in metabolically active, membrane-bound organelles. Science 376:1453–1458.
Woese, C. R., and G. E. Fox. 1977. The concept of cellular evolution. Journal of Molecular Evolution 10:1–6.
Zachar, I. 2022. Closing the energetics gap. Nature Ecology & Evolution.
Zachar, I., and G. Boza. 2020. Endosymbiosis before eukaryotes: mitochondrial establishment in protoeukaryotes. Cellular and Molecular Life Sciences 77:3503–3523.
Zachar, I., and E. Szathmáry. 2017. Breath-giving cooperation: critical review of origin of mitochondria hypotheses. Biology Direct 12:19.
Zachar, I., A. Szilágyi, S. Számadó, and E. Szathmáry. 2018. Farming the mitochondrial ancestor as a model of endosymbiotic establishment by natural selection. Proceedings of the National Academy of Sciences of the United States of America 115:E1504–E1510.
Kapcsolódó cikkek a Qubiten:
Magyar kutató válaszolhatja meg az evolúciókutatás egyik alapkérdését
A Nature Ecology & Evolution-ben megjelent magyar-angol közös eredmény eldöntheti az evolúciókutatás egyik alapvető dilemmáját, vagyis azt, hogy miként alakultak ki a mai élővilág legnagyobb csoportjai, azaz milyenek az evolúciós törzsfa legkorábbi elágazásai.
Egy titokzatos archea vizsgálatával közelebb jutottunk a komplex élet kialakulásának megértéséhez
Mindössze másodszor sikerült laboratóriumi körülmények között asgard archeát tenyészteni – ezek az élőlények az állatok, növények, gombák és más egysejtűek által alkotott eukarióták egyik legközelebbi rokonai. Szöllősi Gergely evolúciógenetikus segítségével jártuk körül ennek jelentőségét.

Hogyan definiáljuk az életet?
Hogyan jöhettek létre az élet építőkövei? Melyek a legegyszerűbb élőlények? A Qubit podcastsorozatában, a Darwin démonaiban Kun Ádám evolúcióbiológus és Mandl Péter bécsi kutatóorvos keresi a válaszokat.